 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 757-761 Rapid Communication Molecular Tagging and Effect Analysis of a New Small Grain Dwarf Gene in Rice LI Xiu-Lan1, WU Cheng2, DENG Xiao-Jian1, 2*, WANG Ping-Rong1, LI Ren-Duan1, YANG Zhi-Rong2* (1. Rice Research Institute, Sichuan Agricultural University, Chengdu
611130, China;
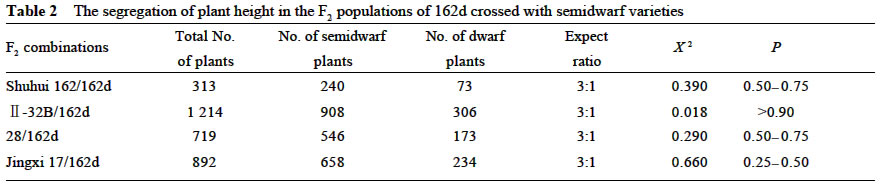
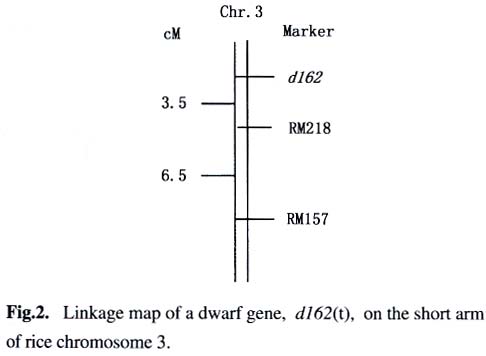
2. College of Life Sciences, Sichuan University, Chengdu 610064, China) Received: 2002-12-13 Accepted: 2003-03-02 Code Number: as03001 Plant height is one of the important agronomic traits of rice. Over higher plant would easily result in plant lodging and output reducing. On the other hand, the dwarf varieties with proper plant height had higher lodging resistance and a greater harvest index, allowing for the increased use of nitrogen fertilizer. Dwarf breeding had made a great breakthrough in the rice breeding. The breeding and extension of excellent dwarf varieties remarkably improved the yield potential of rice. Therefore, the plant height is still one of the focuses in rice genetic research. So far a great deal of study has been made on the dwarf trait of rice, and about 60 dwarf and semidwarf genes have been reported (Kinoshita, 1984; Nagato and Yoshimura, 1998). More than 20 dwarf and semidwarf genes have been located on the rice chromosomes by using methods of classical genetics, and three semidwarf genes (Liang et al, 1994; Maeda et al, 1995; Jiang et al, 2002) and nine dwarf genes (Kishimoto et al, 1992; Yoshimura et al, 1992; Yu et al, 1992; Abenes et al, 1994; Ideta et al, 1994a; 1994b; Ashikari et al, 1996) have been mapped by using molecular markers. Recently, two rice dwarf genes, d1 (Ashikari et al, 1999 ) and d61 (Yamamuro et al, 2000 ), have been cloned, which resulted in dwarf by acting on the signaling pathway of the endohormone. In the present study, a small grain dwarf mutant, designated as "162d" , was identified from a semidwarf variety Shuhui 162, and the mutant gene was mapped with microsatellite markers. Meanwhile, the phenotype effects of the mutant gene were analyzed by using the near isogenic lines. Key words: Oryza sativa ; dwarf gene; gene mapping; microsatellite marker; gene effect analysis 1 MATERIALS AND METHODs 1.1 Plant materials and field experiments 162d was a small grain dwarf mutant identified from a semidwarf variety Shuhui 162 of rice (Oryza sativa L.). 162d was crossed with four semidwarf varieties, Shuhui 162 (indica), II-32B (indica), 02428 (japonica) and Jingxi 17 (japonica). Under natural condition in Chengdu, Sichuan Province, parents and their F1 hybrids and F2 populations were planted, and the plant height was measured one by one at the maturity. 1.2 Construction of mapping population and extraction of total genomic DNA The mapping population consisted of 30 dominant semidwarf individual plants and 96 homologous recessive dwarf individual plants in the F2 population of II-32B/162d. The leaf of each selected individual was used for DNA extraction. At the same time, 10 semidwarf plants and 10 dwarf plants were selected randomly from the mapping population. To construct the DNA pools of dwarf and semidwarf, leaf tissue from all of 10 individual plants within a class were bulked equally for DNA extraction. Total genomic DNA was extracted as described by McCouch et al (1988). 1.3 SSLP analysis SSLP analysis was performed according to Panaud et al (1996), and the amplification products were run in 3.0%-3.5% agarose gels. 1.4 Genetic mapping of the target gene on chromosome region According to the result of SSLP analysis, the band pattern being identical with that of II-32B and 162d was marked 1 and 2, respectively, and the hetero band pattern was marked 3. Then, based on the segregation data of the plant height and molecular markers of the F2 mapping population, the linkage map was constructed with Mapmarker/EXP (version 3.0). The genetic distance (centimorgan, cM) was derived with Kosambi function. 2 RESULTS AND ANALYSES 2.1 Genetic analysis of the dwarf trait of 162d The F1 progenies from 162d crossed with the semi-dwarf varieties, Shuhui 162, II-32B, 02428 and Jingxi 17, were similar to their semidwarf parents in plant height (Table 1). At the same time, plant height of the individual plants distributed bimodally in the F2 populations. The individuals in each F2 population could be easily classified into two groups, semidwarf and dwarf, according to their plant height. The segregation ratio of semidwarf plants to dwarf plants fitted the ratio of 3:1 in the F2 populations (Table 2). These results indicate that the dwarf trait of 162d is controlled by one recessive nuclear gene. 2.2 Molecular marker mapping of the dwarf mutant gene of 162d The F2 population of II-32B/162d was used as the mapping population of the mutant gene. The genomic DNA of parents,II-32B and 162d, were amplified with 204 pairs of primers of microsatellite markers well-distributed on 12 pairs of rice chromosomes. The DNA pools of semidwarf and dwarf were amplified with the microsatellite primers with parental polymorphism, and then the genomic DNA of the 2 individuals amplified with the primer(s) displaying polymorphism between the two pools. As a result, the microsatellite marker, RM218, locating on the short arm of rice chromosome 3, was found to be linked to the dwarf mutant gene of 162d (Fig.1). The genomic DNA of parents,II-32B and 162d, were amplified with microsatellite markers locating nearby RM218. The parental polymorphism of microsatellite marker RM157 was observed, and the genomic DNA of F2 individuals was amplified with RM157. The results suggest that there is linkage relationship between RM157 and the target gene. The linkage map of the dwarf mutant gene was constructed with the segregation data of the plant height and microsatellite markers of the F2 mapping population of II-32B/162d (Fig.2). The results show that the dwarf gene located on the one side of the microsatellite markers, RM218 and RM157, on the short arm of rice chromosome 3, and the genetic distances from the target gene to the markers, RM218 and RM157, are 3.5 cM and 10.0 cM, respectively. This gene, probably being a new dwarf gene in rice, is designated tentatively as d162(t). 2.3 Effect analysis of the mutant gene of 162d
DISCUSSION
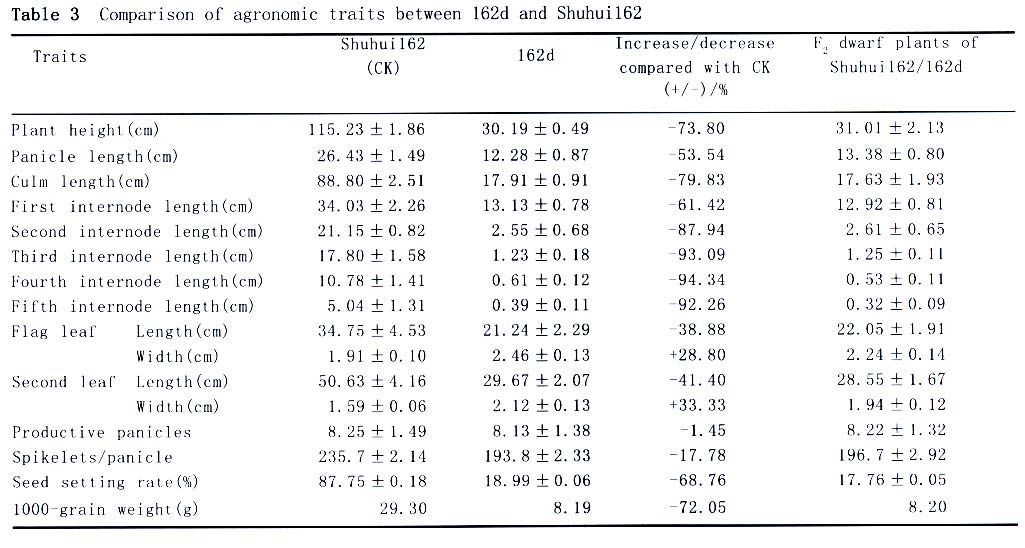
162d was identified from the population of semidwarf variety Shuhui 162, so the genomic DNA of 162d and Shuhui 162 was analyzed with the microsatellite markers well-distributed on the 12 pairs of rice chromosomes (Temnykh et al, 2000). As a result, no polymorphism existing between 162d and shuhui 162 was found among 221 microsatellite loci. So, it can be inferred that 162d derive from the mutation of Shuhui 162, and 162d and Shuhui 162 were a pair of near isogenic lines. During the maturity, 10 individual plants were selected randomly from the populations of 162d and Shuhui 162, respectively, and the productive panicles of the individual plant and other agronomic traits of the main panicle were examined. It was found that 162d had the following characteristics compared with its near isogenic line Shuhui 162 (Table 3; Fig.3): (1) The plant was extremely dwarfed. The plant height and culm length decreased by 73.8% and 79.8%, respectively. The first internode length was shortened 61.4%, and the second, third, fourth and fifth internode length was shortened from 87.9% to 94.3%. (2) The seed became notably smaller. One thousand-grain weight was cut down 72.1%. (3) The leaf was shortened and broadened. The flag leaf and the second leaf were shortened 38.9% and 41.4%, broadened 28.8% and 33.3%, respectively. (4) The filled spikelets per panicle remarkably decreased. The seed setting rate was only 19.0%, which dropped by 68.8%. (5) The productive panicles had no change basically and spikelets per panicle decreased a little (spikelets/panicle decreased by 17.8%), but panicle length was shortened 53.5%. At the same time, the semidwarf and dwarf individual plants in the F2 population of Shuhui 162/162d were observed one by one in the field. It was found that the phenotypes of semidwarf and dwarf plants were very similar to that of Shuhui 162 and 162d, respectively, and there was no small grain individual in the semidwarf plants and no large grain individual in the dwarf plants. Then, 10 individual plants were also selected randomly from the F2 population between small grain dwarf mutant 162d (left) and its near isogenic line Shuhui 162 (right) of Shuhui 162/162d, and their agronomic traits were analyzed by using the above-mentioned method, the same results as that in the field was obtained (Table 3). Therefore, it is considered that the above-mentioned characteristics would result from pleiotropism of the mutant gene of 162d. Based on the above-mentioned results, the phenotype effects of the mutant gene of 162d were summarized as follows: the plant height shortened about three quarters; the grain became extremely small, the 1 000-grain weight reduced about three quarters; the leaves shortened and broadened remarkably; the seed setting rate was very low, less than 20%. Therefore, the mutant gene of 162d can be defined as the small grain dwarf gene of rice. Among about sixty of rice dwarf and semidwarf genes reported so far, eight genes, i.e. d1, d7 , d11, d13, d24(t), d28, d30 andd58, were classified as small grain dwarf genes, but only d1 's phenotype was basically similar to 162d (Kinoshita, 1984; Nagato and Yoshimura,1998; Ashikari et al, 1999). The d1 gene located between C309(S2351) and G1458 near the centromere region of the rice chromosome 5, 0.8 cM and 3.9 cM from C309 and G1458, respectively (Ashikari et al, 1996; 1999; Harushima et al, 1998). In the present study, the microsatellite analysis suggest that the small grain dwarf gene of 162d is not located on the region where d1 gene is located and that the mutant gene is located on the short arm of rice chromosome 3. Therefore, the mutant gene of 162d is considered to be a new small grain dwarf gene in rice and designate tentatively as d162 (t). The rice dwarf mutant carrying the d1 gene, was not only short, one third of normal plant height, but had short, round grains and broad, dark green leaves (Kinoshita, 1984; Ashikari et al, 1999). In the present study, the polymorphisms of 221 microsatellite loci indicate that 162d derived from the mutation of Shuhui 162, and 162d and Shuhui 162 are a pair of near isogenic lines. According to the investigation of agronomic traits of 162d, Shuhui 162 and their F2 individuals, it was known that the d162(t) gene could reduce the plant height and grain size to about a quarter of that of its near isogenic line Shuhui 162, and could shorten and broaden the leaves evidently. So, the phenotype effects of d162(t) gene was greater than that of d1 gene. In addition, the rice dwarf mutant, 162d, had low seed setting rate, only being 19.0%, and being 68.8% lower than that of Shuhui 162. Until now, the rice dwarf mutant with so low seed setting rate have not been reported. The microscope examination showed that the pollen fertility of 162d was similar to that of Shuhui 162. Therefore, the reason resulting in low seed setting rate of 162d is still waiting for deep study. REFERENCES:
(Managing editor: HE Ping) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03001f1.jpg] [as03001f2.jpg] [as03001t3.jpg] [as03001t2.jpg] [as03001f3.jpg] [as03001t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}