 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 770-773 Rapid Communication Agroinoculation as a Simple Way to Deliver a Tobacco Mosaic Virus-Based Expression Vector JIA Hong-Ge1 , PANG Yong-Qi1, 2 , FANG Rong-Xiang1 * (1. Laboratory of Plant Biotechnology, Institute of Microbiology, The
Chinese Academy of Sciences, Beijing 100080, China; Received: 2002-12-02 Accepted: 2003-03-11 Code Number: as03004 As an expression vector with characteristics of relatively high stability and high-level production of heterologous proteins, the tobacco mosaic virus (TMV)-based 30B vector has been available for the experimental use worldwide (Yusibov et al, 1997; Shivprasad et al, 1999; Nemchinov et al, 2000). In a conventional procedure, however, a step of in vitro transcription is required to produce infectious viral RNA prior to inoculation of host plants. This troublesome and expensive process has hindered the use of the 30B vector to some extent. To conquer such disadvantage, we have adopted a technique called agroinoculation in which the cDNA copy of a viral genome, herein 30B, is inserted into the T-DNA under the control of the cauliflower mosaic virus (CaMV) 35S promoter in such a way that it can be easily delivered into plant cells and initiate an infection. In order to test the agroinoculation method conveniently, the gfp gene for the jellyfish green fluorescent protein (GFP) was used as a reporter gene and inserted into the cloning sites of the viral vector to construct plasmid p35S-30B:: GFP. The agrobacteria harboring the modified vector were simply pressure-infiltrated into a leaf using a needle-less syringe. In this paper, we report high-level expression of GFP in systemic leaves up to 5.2% of total soluble leaf protein, demonstrating that agroinoculation can be used as an efficient and simple way to express foreign proteins in plants from the 30B vector. Key words: tobacco mosaic virus; agroinoculation; gene expression 1 MATERIALS AND METHODS 1.1 Materials
Nicotiana benthamiana was used as the host plant. Agrobacterium tumefaciens strain EHA 105, wild-type TMV (common strain) and rabbit anti-TMV serum are kept in our laboratory. Escherichia coli-expressed GFP and rabbit antiserum against GFP were previously prepared in our laboratory. 1.2 Plasmid construction All molecular manipulations were carried out following standard procedures (Sambrook et al, 2001). The cDNA fragment of the TMV RNA prepared from the 30B vector (Shivprasad et al, 1999) was inserted into the cloning sites between the 35S promoter and terminator of pCassKS (Lei et al, 2001), a derivative of pCass2 (Shi et al, 1997). The DNA fragment containing the 30B cDNA with the upstream 35S promoter and the downstream 35S terminator was cloned into pCambia 1 300 to obtain plasmid p35S-30B. The gfp gene was PCR-amplified from pBIN 35S-mgfp5-ER (Haseloff et al, 1997) using a forward primer P1 (5'-attaattaatgaagacta atctttttctc-3') and a reverse primer P2 (5'-actcgagttaaagctcat-catgtttgta-3'), and inserted into the cloning sites of 30B cDNA to form p35S-30B::GFP (Fig.1, A, B), which was then introduced into Agrobacterium strain EHA 105 by electroporation. 1.3 Agroinoculation N. benthamiana was grown in pots at 25 °C in a growth chamber under 16 h light/8 h dark cycles. Agrobacterium EHA 105 harboring p35S-30B::GFP, abbreviated to Agro 35S-30B::GFP, was grown overnight at 28 °C in 3.5 mL of LB containing kanamycin (25 µg/mL) and rifampicin (15 µg/mL). Then 1 mL of the overnight culture was diluted with 50 mL of LB containing the antibiotics, 10 mmol/L MES (pH 5.6) and 20 µmol /L acetosyringone. The culture was incubated overnight at 28 °C, with shaking at 300 r/min. Agrobacterium cells were collected by centrifugation at 3 000 r/min, resuspended in solution MMA (10 mmol/L MgCl2, 10 mmol/L MES (pH 5.6), 100 µmol/L acetosyringone) , adjusted to a final OD600 of 0.6 and left at room temperature for 3 h without shaking, and then pressure-infiltrated into a leaf of a N. benthamiana plant using a 2-mL syringe without needle. 1.4 Detection of GFP
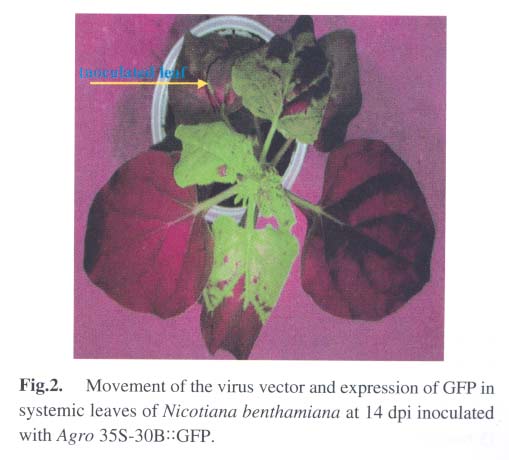
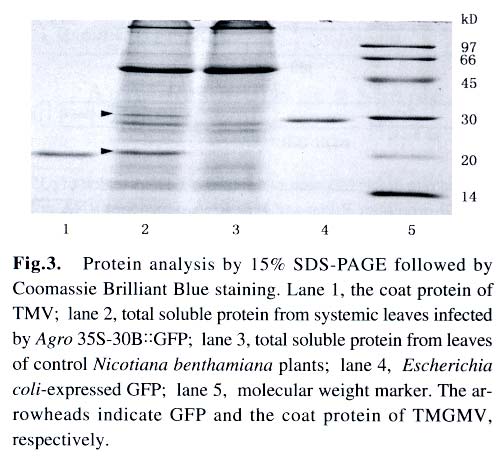
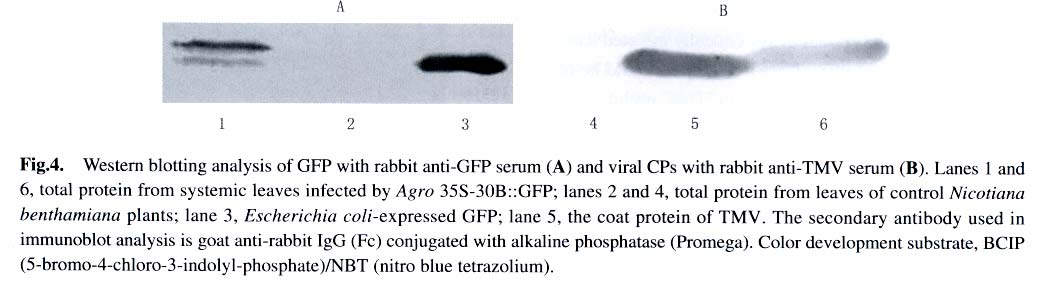
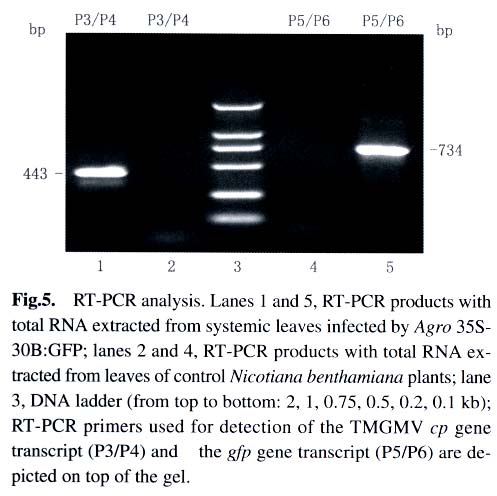
Under illumination of a 100 W hand-held long-wave ultraviolet lamp (UV Products, model B 100AP), the fluores-cence of GFP in intact plants was visualized or photographed using a digital camera Nikon Coolpi × 995. 1.5 Analysis of GFP and viral coat protein (CP) in systemically infected leaves For plant protein analysis, 100 mg of leaves showing green fluorescence was ground into powder in liquid nitrogen. The homogenate (100 µL) was mixed with equal volume of 2 × loading buffer and boiled for 3 min. After centrifugation at 14 000 r/min for 5 min, the supernatant was loaded for SDS-PAGE, followed by Coomassie Brilliant Blue R-250 staining and Western blotting analysis. 1.6 RT-PCR analysis of the gfp and cp gene transcripts in systemically infected leaves Total RNA was isolated from systemic leaves with the guanidine thiocyanate method. First-strand cDNA was synthesized using avian myeloblastosis virus (AMV) Reverse Transcriptase (Promega) according to the manufacturer's instruction. A fraction of the first-strand cDNA was used as a template in the following PCR procedures. Primers P3 (5'-tgaactggttcgtggaactggc-3') and P4 (5'-ggccgctacccgcggtta-3') were used for analysis of the cp gene transcript. Primers P5 (5'-tggatccaacaatgagtaaaggaga-agaacttttc-3') and P6 (5'-gagctcttatttgtatagttcatccatgcc-3') were used for detection of the gfp gene transcript. VentR® DNA Polymerase (Biolabs) was used for all the PCR amplifications with the following cycling conditions: 94 °C for 3 min; 35 cycles at 94 °C for 45 s, 54 °C for 1 min, 72 °C for 1 min; followed by a final incubation at 72 °C for 10 min. The RT-PCR products were resolved by electrophoresis in a 1.2% agarose gel. 2 RESULTS AND DISCUSSION Under UV illumination, green fluorescent spots could be clearly visualized in agroinoculated leaves of N. benthamiana at 5 d post-inoculation (dpi). Diffused green fluorescence of GFP could be detected in upper uninoculated leaves at approximately 9-11 dpi (Fig.2). However, when the extract of systemically infected leaves was used as an inoculum, fluorescent spots could be observed in inoculated leaves at 3 dpi. This implies that a period of about 2 d may be required for in vivo transcription of viral RNA from T-DNA, replication of the nascent viral RNA by RNA-dependent RNA polymerase (RdRp) encoded by the viral genome, and the accumulation of sufficient GFP. Among 40 inoculated plants, 37 exhibited the systemic infection of virus with expression of GFP. GFP produced in Agro 35S-30B::GFP-inoculated plants was readily detectable when the total plant protein was analyzed by SDS-PAGE followed by Coomassie Brilliant Blue staining (Fig.3). The amount of GFP in total soluble leaf protein was estimated to be 5.2% of total soluble leaf protein based on the densitometer scanning profile of the stained gel using the software TotalLab v1.00 (data not shown). However, two forms of GFP with different molecular weights appeared in the Western blotting analysis (Fig.4A), similar to those observed in transgenic plants transformed by pBIN 35S-mgfp5-ER (data not shown). It is possible that the GFP band with higher molecular weight still retains the signal peptide derived from a chitinase of Arabidopsis thaliana owing to insufficient proteolytic processing in N. benthamiana. Since TMV and tobacco mild green mosaic virus (TMGMV) are serologically related, the anti-TMV serum was used to detect the coat protein of TMGMV in the western blotting analysis (Fig.4B). The RT-PCR results for the gfp and cp gene transcripts provided further evidence for systemic spread of the virus-based vector with the foreign sequence (Fig.5). In order to study posttranscriptional gene silencing and gene function conveniently, several viral vectors have been modified for agroinoculation, such as potato virus X (Jones et al, 1999) and tobacco rattle virus (TRV) (Ratcliff et al, 2001; Liu et al, 2002). The TRV-based vector and the TMV 30B vector were used as powerful tools to study genes involved in N -mediated resistance to TMV by the agroinoculation method (Liu et al, 2002). More recently, the TMV 30B vector was used in a virus-induced gene silencing system to reveal the dynamics of the silencing of both the ChlH gene encoding the H subunit of magnesium chelatase and the TMV vector per se in N. benthamiana (Hiriart et al, 2003). On the other hand, DNA shuffling was employed to improve the movement and host range properties of the 30B vector (Toth et al, 2002). However, in these two researches, 30B was first transcribed in vitro to produce infectious RNA prior to inoculation of host. These studies indicate that the use and further modifications of the plant virus vectors, including the improvement of the delivery system for the TMV 30B vector as described in this report, will facilitate research in many fields of biological sciences. In plants, heterologous genes can be expressed stably or transiently. Stable transformation of plants is a time-consuming process and some plant species are not amenable to transformation. Furthermore, transgene integration in plant chromosomes and the copy number of foreign DNA sequences may affect the expression level of the foreign proteins. In contrast, virus-mediated transient expression offers a number of advantages, including a short cycle time, ease of scale-up, and a generally wide host range that allows expression of a gene in different plant species using the same vector construct. Although viral vectors have certain advantages over transgenic plants, they are generally not suitable to express large proteins owing to the virus packaging constraints. Nevertheless, the virus-based vectors are playing important roles in commercial biotechnology, therapeutic treatments and laboratory research (Johnson et al, 1997; Yusibov et al, 1999; Fooks, 2000). ACKNOWLEDGEMENTS We thank Drs. William DAWSON, Jim HASELOFF, DING Shou-Wei, Richard JEFFERSON for providing 30B, pBIN35S-mgfp5-ER , pCass2 and pCambia 1300, respectively. REFERENCES
(Managing editor: ZHAO Li-Hui) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03004f4.jpg] [as03004f2.jpg] [as03004f3.jpg] [as03004f5.jpg] [as03004f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}