 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
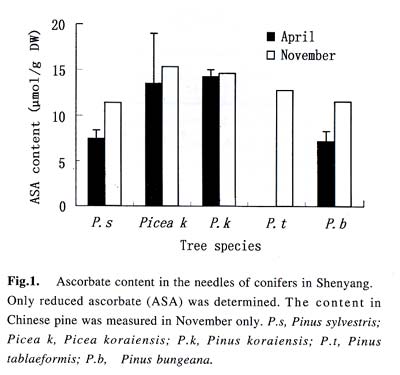
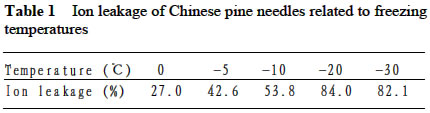
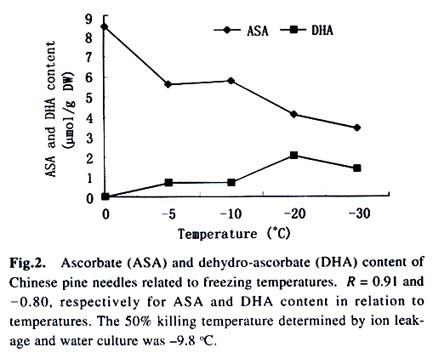
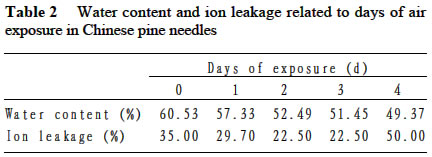
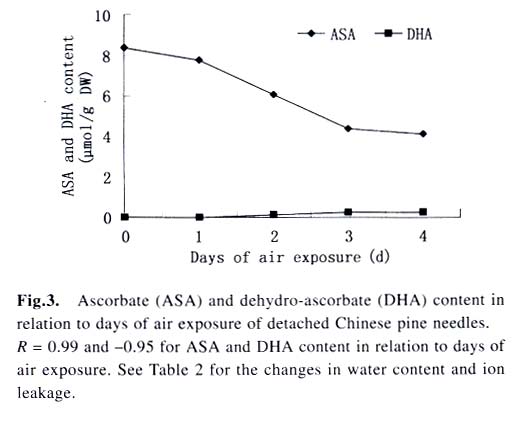
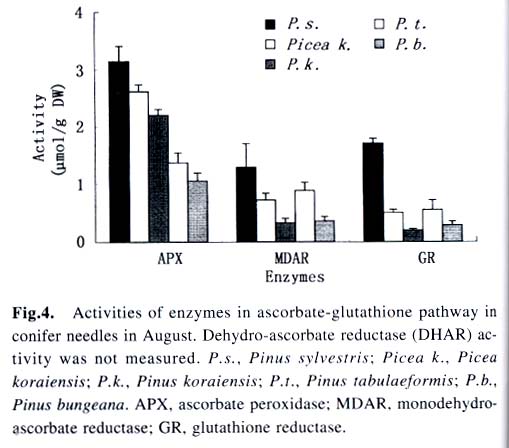
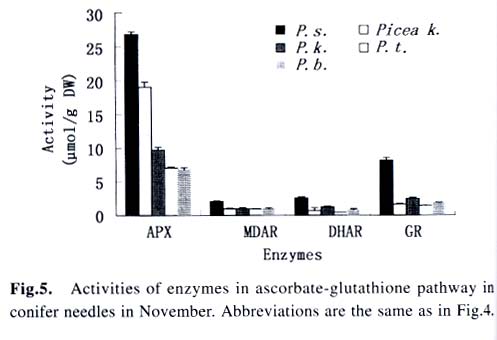
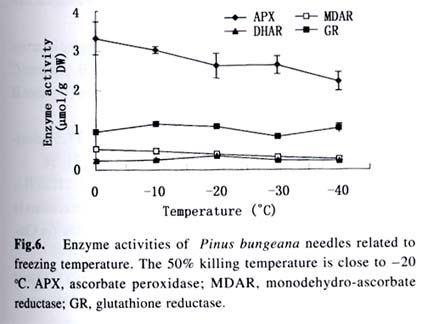
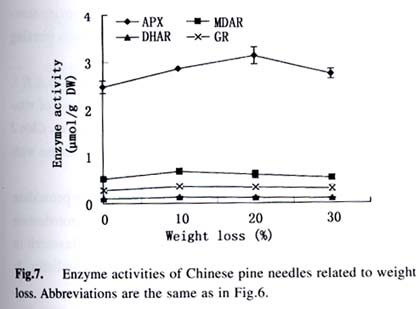
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 795-801 Environmental Stresses and Redox Status of Ascorbate JIN Yue-Hua1, TAO Da-Li1*, HAO Zhan-Qing1, YE Ji, DU Ying-Jun1, LIU Hai-Ling1, ZHOU Yong-Bin2 (1. Key Laboratory of Terrestrial Ecological Process Institute of Applied Ecology, The Chinese Academy of Sciences, Shenyang 110016, China; Received: 2002-07-16 Accepted: 2003-01-09 Code Number: as03007 ABSTRACT To investigate the effects of environmental stresses on ascorbic acid content and its redox status, the effects of freezing and drought on ascorbate and dehydro-ascorbate content and activities of four enzymes involved in the ascorbate-glutathione cycle in some conifers were studied. The results showed that both freezing and drought induced the decrease in ascorbate content and the increase in dehydro-ascorbate content. The activities of ascorbate peroxidase (APX) and monodehydro-ascorbate reductase (MDAR) were decreased by freezing stress. At the beginning of exposure to air, water loss from detached needles induced the increase in the activities of APX and MDAR. Further water loss turned to decrease the APX and MDAR activities. The activities of dehydro-ascorbate reductase (DHAR) and glutathione reductase (GR) were not sensitive to changes in temperature and water content of the needles. It is concluded that moderate tempera- ture or water stresses may induce the acclimation and increase in the ability of the H2O2 scavenging system, while strong stresses decrease the ability and induce injury of plant tissues. Correlation between ascorbate content and activities of related enzymes and cold tolerance of conifers were also reported. Key words: freezing; drought; ascorbate; ascorbate-glutathione pathway Various environmental stresses induce the production of superoxide (O2 -.) that can be scavenged by a specific scavenger superoxide dismutase (SOD) and other scavengers. SOD dismutase O2 -. to O2 and reactive oxygen species H2O2. According to Asada and Takahashi (1987), all H2O2 in chloroplasts is derived from the dismutation of O2 -. . Photosynthesis will be inhibited within seconds if H2O2 is not removed timely (Asada and Takahashi, 1987). There is an ascorbate-glutathione metabolic pathway called Halliwell-Asada cycle (van Montagu and Inzé,1992) that is responsible for the scavenging of H2O2. This cycle was originally found in chloroplast. But later on, it became known that it also operated in other parts of cells and tissues. SOD cooperates with this cycle in scavenging reactive oxygen species. An increase in SOD activity alone may induce accumulation of H2O2 that is not timely removed by ascorbate-glutathione cycle (van Montagu and In z é,1992). We have shown that during cold acclimation of Scots pine (Pinus sylvestris), ascorbate content and the activities of enzymes involved in the ascorbate-glutathione cycle increased in parallel to SOD activity. These increases were suggested to have the effect in protecting pine needles from photoinhibition (Krivosheeva et al, 1996) and freezing stress (Tao et al,1998). Ascorbate (ASA) plays a central role in ascorbate-glutathione cycle where it is oxidized by ascorbate peroxidase (APX) to the monodehydro-ascorbate (MDA) radical and then to dehydro-ascorbate (DHA). MDA is reduced by monodehydro-ascorbate reductase (MDAR) with NAD (P) H as the reductant and DHA is reduced by dehydroascorbate reductase (DHAR) with glutathione to regenerate ascorbate (Asada, 1994). Ascorbate also participates in the direct removal of O2 -. , regeneration of tocopherol and in the reduction of violaxanthin to antheraxanthin and zeaxanthin in the xanthophyll cycle (Foyer, 1993). However, our knowledge of ascorbate is still scarce. Even the biosynthetic pathway of ASA in higher plants is still not clear (Wheeler et al, 1998). Maximum ascorbate content in needles of Korean pine (Pinus koraiensis) was detected in midwinter in Changbai Mountain and its content in sun-exposed needles was three times to those under shading (Tao et al, 1992). Cold acclimation induced higher activities of all the enzymes in ascorbateglutathine cycle (Krivosheeva et al, 1996, Tao et al, 1998). Here we present some data to show the effects of non-injurious and injurious stresses on ASA content and activities of its oxido-reduction enzymes. The proportion of DHA in the ascorbatedehydroascorbate pool was increased from 9% to about 50% in the process of cold acclimation of Scots pine seedlings that was related to the high activity of APX and low activities of dehydro-ascorbate reductase (DHAR) and monodehydroascorbate radical reductase (MDAR) (Tao et al, 1998). This suggests that the regeneration of ascorbate might be a limiting factor in enhancing the cold tolerance of Scots pine and the shortage of reduced ascorbate might be a factor in inducing injury. It is also interesting to know if there is any relation between cold tolerance and ASA content and activities of the four enzymes in the ascorbate-glutathione cycle mentioned above in other conifer species. Here we show that the activities of the four enzymes are related to the already known cold tolerance of five conifers in Shenyang (Jin et al, 1989). The imbalance between consumption and regeneration of ascorbate and, accordingly, the imbalance between the activities of APX versus MDAR and DHAR due to drought and freezing stresses in the conifer needles are also reported. 1 MATERIALS AND METHODS 1.1 Plant materials Adult trees of conifers grown in the yard of the Institute of Applied Ecology, The Chinese Academy of Sciences, were used. ASA content and related enzyme activities were compared among five species, namely Mongolian pine (Pinus sylvestris var. mongolica) native to Da Hinggan Mountains, Korean spruce ( Picea koraiensis Nakai) native to Da Hinggan Mountains, Xiao Hinggan Mountains and Changbai Mountain , Korean pine native to Xiao Hinggan Mountains and Changbai Mountain but not to Da Hinggan Mountains, Chinese pine (Pinus tabulaeformis Carr.) native to Shenyang and Bunge pine (Pinus bungeana Zucc.) native to Beijing but marginally surviving the cold winter in Shenyang. According to their native ranges of distribution, it is apparent that the first two species are the most cold tolerant. Korean pine is the next. Bunge pine is the most vulnerable and its foliage was injured in severe winter in Shenyang. The difference in their cold hardiness was also evidenced in the deep freezer test: In midwinter, the 50% killing temperature was –33 oC for Bunge pine and –70 oC for Chinese pine. The other three conifers all survived at –70 oC without any injury (Jin et al, 1989). Chinese pine and Bunge pine were used for freezing and drought stress experiments. Current year needles were collected in the morning and immediately used for experiments. In the experiments on drought, parallel samples were taken and dried at 80 oC to measure the water content before the needles were air-dried in the laboratory. 1.2 Freezing stress Pine needles were cooled in a thermos placed in a deep freezer. The junction of a thermocouple was attached to needles to ensure the temperature of needles per se was monitored. The temperature was lowered stepwise by 5 oC for 1 h to –10 oC and then 10 oC for 1 h till –30 oC. At the end of each step, a sample was taken out and thawed in another thermos containing crushed ice and water to ensure 0 oC and put in a refrigerator until next morning. 1.3 Drought stress One gram (for enzymes assay) or 0.5 g (for ascorbate assay) freshly detached needles were weighed and then exposed to the air on a bench in the laboratory for various periods and the weight losses were recorded. Parallel samples were exposed to the same environment and used to measure changes in water content and viability. The experiments were done twice, in August and in December. 1.4 Viability test After treatments with all stress types and strengths, samples were tested for their viability by measuring their ion leakage and behavior in water culture. For ion leakage measurement, the needles were cut into pieces, soaked in distilled water and shaken for 5 h. Electric conductivity of the water was then measured. After the samples were killed at 80 oC in a water bath for 20 min and cooled to room temperature, conductivity was measured again and the percentage of ion leakage was calculated. For water culture, the basal parts of needles were soaked in water to observe the change in appearance. The healthy needles retained their normal green color and turgidity for a few months while the injured ones wilted and became discolored in a few days. The 50% survival was judged according to the ratio between total lengths of green and brown needles. In our long-term experiences, 50% ion leakage was quite close to 50% survival in water culture. 1.5 Ascorbate and dehydro-ascorbate assay High performance liquid chromatography (Liu et al, 1994) was used for ascorbate assay before and after the dehydroascorbate in the extract was reduced to ascorbate by added glutathione. Half a gram of needles was ground in liquid nitrogen. After thawing, 4.5 mL (pH 5.6) phosphate buffer was added. After filtering through nylon cloth, the homogenate was centrifuged at 16 000 g and 4 oC for 20 min. The supernatant was used for assaying enzyme activities. For ASA assay, 0.9 mL extract was mixed with 0.1 mL buffer. For the ASA+DHAassay, 0.9 mL of the extract was mixed with 0.1 mL GSH 0.1 mol/L in phosphate buffer (pH 7.6). The mixture was kept at room temperature for 15 min to ensure that DHA in the extract was fully reduced to ASA by GSH. 1.6 Enzyme extraction One gram of needles was ground in a mortar in liquid nitrogen. Right after the product was thawed in a refrigerator, the crude enzymes were extracted with 3.5 mL phosphate buffer containing 2% PVP, EDTA 1 mmol/L and ascorbate (sodium salt, freshly prepared) 1 mmol/L and 0.25% Triton X-100 at pH 7.6. Then 1.5 mL of saturated ammonium sulfate in the samebuffer was added and stirred well to minimize the interference of resin that would otherwise inhibit the APX activity (Krivosheeva et al, 1996). After filtering through nylon cloth, the filtrate was centrifuged for 20 min at 16 000 g and 4 oC. The supernatant was transferred into vials and then kept on ice and the enzyme activities were assayed immediately. 1.7 Enzyme activity assay The enzyme assays followed Krivosheeva et al (1996). For APX assay, 1 mL reaction mixture containing 0.83 mL of 0.5 mmol/L ascorbate in phosphate buffer (pH 7.0), 0.13 mL of 2 mmol/L H2O2, both of which were freshly prepared and 0.04 mL of crude enzyme was used. The ascorbate consumption was monitored by the reduction of absorbance at 290 nm taking 2.8 (mmol/ L)-1 cm-1 as the absorption coefficient (Nakano and Asada, 1981). For the MDAR assay, a reaction mixture containing 0.9 mL of 2 mmol/L ASA in phosphate buffer (pH 7.0), 0.04 mL of ASA oxidase (2 units) in phosphate buffer (pH 5.6), 0.03 mL of 2 mmol/L NADPH in buffer (pH 7.6) and 0.03 mL crude enzyme was used. The consumption of NADPH was monitored by the reduction of absorbance at 340 nm taking 6.2 (mmol /L)-1cm-1 as the absorbance coefficient (Krivosheeva et al, 1996). For the DHAR assay, a reaction mixture containing buffer (pH 7.0) 0.7 mL, reduced glutathione (GSH) 20 mmol/L 0.1 mL in buffer (pH 7.0), 2 mmol/L DHA 0.1 mL and crude enzyme 0.1 mL was used. DHA was freshly prepared and kept on ice till adding to the reaction mixture in the cuvette to prevent its fast oxidation at room temperature. The reduction of DHA to ASA was monitored by the increase in absorbance at 265 nm taking 14 (mmol/ L)-1 cm-1 as the absorbance coefficient (Krivosheeva et al, 1996). For glutathione reductase (GR) assay, the reaction mixture containing 0.86 mL of 1 mmol/L oxidized glutathione (GSSG), 0.1 mL of 2 mmol/L NADPH in phosphate buffer (pH 7.6) and 0.04 mL crude enzyme was used. The consumption of NADPH was monitored for MDAR assay. All the enzyme reactions were monitored for 1 min in the cuvette of a Shimazu UV-1601 spectrophotometer. NADPH, DHA, GSSG and ascorbate oxidase were bought from Sigma. All the enzyme assays were repeated three times. 2 RESULTS 2.1 Ascorbate content in needles of conifers in Shenyang In April, only ascorbate in the reduced form was determined with replications but Chinese pine was not measured. In November, total ascorbate and two forms of ascorbate were all determined. The dehydro-ascorbate contents were found to be all below the limit of detection of the method used. Results (Fig. 1) showed the general trend that the more cold-tolerant the species, the more ascorbate content in the needles it had. Mongolian pine was the only exception. The ASA content was higher in November than that in April. 2.2 Ascorbate status affected by environmental stresses In freezing test of Chinese pine needles carried out in September, water culture showed that most needles sustained slight injury indicated by tip discoloration after being frozen to -5 °C. The injury increased at –10 oC, indicating by discoloration of a little more than 1/2 of the length in all needles. This is quite consistent with the 53.8% ion leakage (Table 1). The calculated half-killing temperature was –9.8 o C. The ascorbate content decreased with lowering of freezing temperatures with r = 0.90 while the dehydro-ascorbate content showed an increase (Fig. 2). Similar and better results were got from drought experiment done in August. The exposure of needles to the air in the laboratory caused gradual water loss (Table 2). Close correlation (r = 0.964) between ascorbate content (Fig. 3) and water content (Table 2) of the needles was shown. The water culture at room temperature showed normal green and turgid appearance till 4 d of exposure when 18.4% weight or 30% water content were lost. The ion leakage increased sharply to 50% and 6 out of 12 needles in water culture showed discoloration. The decrease in ASA content was detected after 1d of exposure and DHA became detectable after 2 d exposure and then kept increasing. In the drought experiment repeated in December, similar results were obtained. The only difference was that DHA could hardly be detected (Data not shown). 2.3 Activities of enzymes in ascorbate-glutathione cycle in five conifers The lower the temperature a tree species can tolerate (Jin et al, 1989), the higher the enzyme activities in the needles. This trend was most apparent for APX. The activity of MDAR was about half that of APX in August (Fig. 4). In November, when the trees had been cold-acclimated to some extent, the activities of all the four enzymes were higher than those in August (Fig. 5). However, the extent of increase in activities was quite different among the enzymes studied. The activity of APX in November was seven times higher than that in August while the activity of other enzymes increased much less. The total activity of MDAR +DHAR in November was only 17% of that of APX. The disparity between the abilities to use and to regenerate ascorbate was much enlarged. This is consistent with the results of artificial cold acclimation of Scots pine (Krivosheeva et al, 1996; Tao et al, 1998). 2.4 Enzyme activities affected by stresses In the freezing experiment with Bunge pine that had a 50% killing temperature of about -20 oC, the activity of APX and MDAR showed an apparent decrease with lowered freezing temperatures (Fig. 6). The variations in activity of DHAR and GR were less clear. In air exposure experiment done on a clear and hot day in August, the 30% weight loss occurred within one day. Visible injury symptoms—wilt, twisting and loss of brightness on the surface of the needles was evident when more than 20% of the weight was lost. The water content of Chinese pine needles decreased by about one half and the ion leakage was a little more than 50% at the end of the exposure (Data not shown). The enzyme activities increased first and then turned to decrease. The APX activity turned to decrease later than the other three enzymes (Fig. 7). Whatever the stress was, APX activity was consistently much higher than the sum of DHAR plus MDAR activities. In the drought experiment repeated in December, similar results were obtained (Data not shown). 3 DISCUSSION 3.1 Variations in ascorbate and dehydro-ascorbate content We have reported that ASA content in Korean pine needles showed seasonal variation with its maximum in midwinter in Changbai Mountain (Tao et al, 1992). ASA content of Scots pine showed increase during acclimation to artificial cold environment (Krivosheeva et al, 1996; Tao et al, 1998). Here a general trend was that the cold-tolerant conifer tree species had higher ASA content in their needles (Fig.1). These results suggest again that ASA played an important role in cold tolerance of trees. Mongolian pine was the only exception and will be discussed later. Without environmental stress, DHA content was lower than the limit of detection of the method we used. However, when freezing or drought stress was exerted to the needles, DHA became detectable and the DHA content was increased with further exposure to the stresses (Figs. 2, 3), indicating that oxidative stress was a common factor for both cold and drought stresses and the injuries caused by water and cold stresses were related to the over-oxidation or insufficient regeneration of ascorbate. The consumption of ASA and the presence of DHA may be used as indexes of oxidative stress. However, the amount of DHA detected could account for all the consumption of ASA (Figs. 2, 3) because DHA is unstable and may be converted to tartrate, oxalate and other compounds (Foyer, 1993). 3.2 Variations in the enzyme activities 3.2.1 Enzyme activities in relation to cold tolerance of conifers The good correlation between cold tolerance of conifer species and APX activity was shown even in August, a summer month (Fig.4). In November, the difference in APX activity among tree species was larger. Special attention should be paid to the extra-high activity of APX in Mongolian pine. Figure 1 suggested that the low ASA content in Mongolian pine might be compensated by its fast turnover catalyzed by the very high activities of APX (Figs. 4, 5) and enzymes involved in the regeneration of ascorbate. Other enzymes all showed their highest activities in Mongolian pine. 3.2.2 Enzyme activities affected by stresses Both APX and MDAR activities decreased with lowering freezing temperature (Fig. 6). In addition to the effect of freezing on ASA (Fig. 2), it seemed that freezing partly inactivated MDAR. As a result, DHA was accumulated. According to Asada and Takahashi (1987), APX needs the protection by reduced ASA. If the ASA content is lower than 20 µmol/L, APX will lose its activity. It is supposed that the decrease in APX activity and the freezing injury may be related to the inadequate regeneration of ASA via DHA reduction. The idea that freezing injury is related to the disturbance in the H2O2-scavenging system is consistent with the report that the collapse of H2O2 scavenging system is associated with freezing injury in de-acclimated apple flower buds (Kuroda et al, 1992). In contrast to the freezing experiment, in the air exposure experiment the enzyme activities increased in the early stages and then turned to decrease (Fig. 7). The increase in early stages implicated some effect of acclimation to drought. It is interesting to note that the environment was quite harsh to the needles. The needles used in the experiment had been detached from trees and no longer supplied with water, but the enzymes, especially APX, could still increase its activity. The short-term air exposure did not incur injury but induced drought acclimation within hours. Such effect of acclimation was hardly seen in freezing experiment where the temperature was decreased at 5-10 oC/h but occurred much more slowly (Krivosheeva et al, 1996). The fast drought-acclimation is reasonable because drought has been more often and comes faster than the seasonal winter cold. 3.2.3 The imbalance between activities of APX and MDAR + DHAR APX was constitutively high and very sensitive to the environmental changes whether the change led to acclimation or injury, but MDAR and DHAR always responded slowly. APX activity could be used as a sensitive indicator for the stress tolerance of tree species, for acclimation and for the injury caused by environmental stresses. In any case, the activity of APX in oxidizing ASA is much higher than the activity of MDAR and DHAR in regenerating ASA. According to Miyake and Asada (1992) and Miyake et al (1998), MDA, the primary product of ASA oxidation, can be reduced back to ASA by cell components other than MDAR such as ferredoxin and in the case where there is no MDA, the flavoenzymes such as MDAR and GR will donate electrons to di-oxygen instead of MDA, with superoxide radical as the primary product. This implicated that too high an activity of MDAR and GR and shortage of MDA radical may be harmful to the cells by generating more superoxide radicals. 3.2.4 Distinct types of responses to environmental stresses Plants may respond to different strengths of environmental stress in distinct ways. When the stress is too weak to affect the plant, there will be no response. When the severity of the stress is within the tolerable range, the signal will be detected by sensitive plant and the plant will timely mobilize its defense system and be acclimated to tolerate the stress severer than it currently experiencing. When the stress is too strong or comes too fast, the plant will not be able to respond to acclimate to it, the defense system will be adversely affected and the plant will be injured. The sensitivities of different components of the defense system are often different so that they may neither respond to the environmental variation synchronously nor in the same direction as shown in our drought experiment (Fig. 7), when the weight loss increased from 20% to 30%, the activity of APX went up but the activity of MDAR went down. These complexities should be considered in research on the responses of plant to environmental changes. REFERENCES
(Managing editor: HAN Ya-Qin) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03007f3.jpg] [as03007f1.jpg] [as03007t2.jpg] [as03007f4.jpg] [as03007f2.jpg] [as03007f7.jpg] [as03007f5.jpg] [as03007f6.jpg] [as03007t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}