 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 809-814 Different Responses of Camptothecin and 10-Hydroxycamptothecin to Heat Shock in Camptotheca acuminata Seedlings ZU Yuan-Gang*, TANG Zhong-Hua, YU Jing-Hua, LIU Shi-Gang, WANG Wei, GUO Xiao-Rui (Key Laboratory of Forest Plant Ecology, Ministry of Education, Northeast Forestry University,
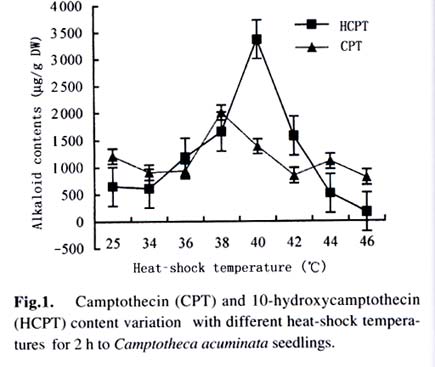
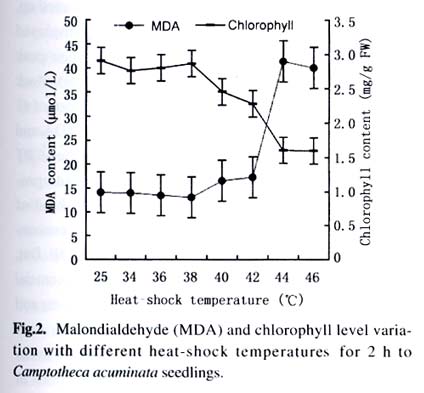
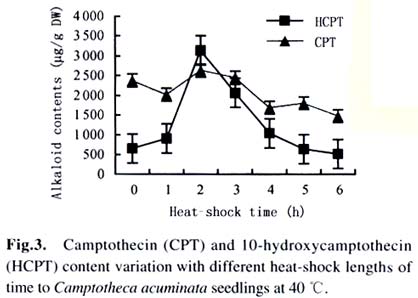
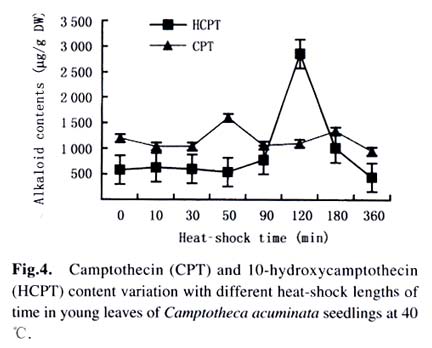
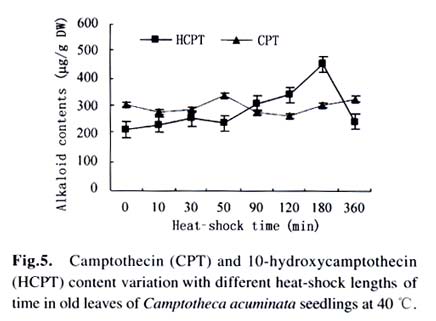
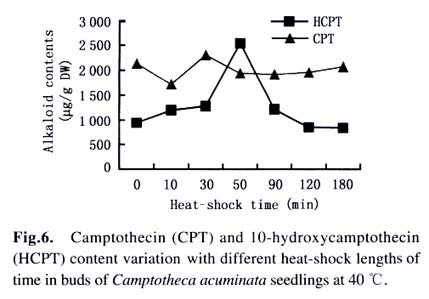
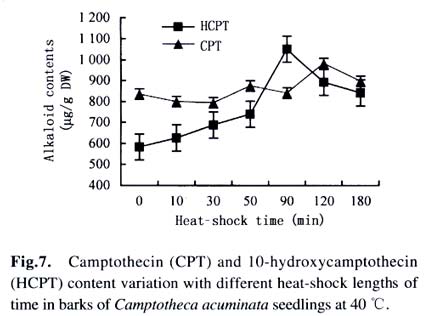
Harbin 150040, China) Received: 2002-12-02 Accepted: 2003-02-12 Code Number: as03009 ABSTRACT Plants interact with their environment by producing a diverse array of secondary metabolites, one of which is alkaloid. In this study, alkaloids, including camptothecin (CPT) and 10-hydroxycamptothecin (HCPT), malondialdehyde (MDA) and chlorophyll contents were measured during heat shock in seedlings of Camptotheca acuminata Decaisne unique to China. Responses of different tissues, including young leaves, old leaves, buds and barks, to heat shock were examined in alkaloid changes. CPT and HCPT concentrations reached their peak values separately at 38 °C and 40 °C, which were below the lethal heat-shock temperature indicated by MDA and chlorophyll, and their great changes took place in young leaves. These results indicated that CPT and HCPT were involved in the C. acuminata resistance against heat shock from its environment. Furthermore, plant rigidly observed the cost-benefit principle and mobilized and allocated limited alkaloid sources to young and reproductive tissues preferentially. In addition, HCPT displayed well-regulated changes during incubation at sublethal temperature, and this indicated that HCPT might play a more positively defensive role in enhancement of plant thermotolerance than CPT does. Key words: Camptotheca acuminata; camptothecin; 10-hydroxycamptothecin; response; heat shock Species-specific secondary metabolites are key players in the integral interaction between plants and their biotic and abiotic environment (Chen et al, 2000). Alkaloids belong to a broad category of secondary metabolites and this class of molecule has traditionally been of interest due to the pronounced and various physiological activities in animals and humans (Kutchan, 2001). With the development of more thorough studies, a picture has now begun to emerge that alkaloids have important ecochemical functions in the defense of plants against pathogenic organisms and herbivore (Rosenthal and Berenbaum, 1991). Damage to leaves, such as browsing herbivores cause, dramatically increases de novo nicotine biosynthesis, the allometricall determined set points and whole-plant nicotine accumulation 2-fold to 10-fold (Baldwin, 2001). Exogenous application of the plant signal molecule MeJA (methyl jasmonate) induces the accumulation of monomeric alkaloids in Catharanthus seedlings (Aerts et al, 1994; Der Fits and Memelink, 2000). Treatment with pathogens induced the increase of phenylpropanoids in soybean. Wounding elicits a massive metabolic commitment to nicotine production, a potent direct defense, which is produced and distributed throughout the plant in a manner that optimizes plant fitness (Baldwin et al, 1994; Baldwin, 1999). Temperature is one of the most active environmental factors and affects all plant metabolic activities (Luthe et al, 2001). The heat-shock response is a highly conserved reaction of cells and organisms to elevated temperatures, because it is often chaperoned by drought, osmotic and oxidative stresses (Schoffl et al, 1998). So, a higher level of thermotolerance is of utmost importance for plants to cope with external stresses. At the molecular level, activation of this response is a transient reprogramming synthesis of HSPs (heat-shock proteins), which allows resumption of normal cellular and physiological activities (Welch, 1992). But little is known about the interaction between heat shock and alkaloid biosynthesis and accumulation. Camptotheca acuminata is uniquely distributed in South China, especially concentrating in Sichuan Province and Guizhou Province. It is fond of heat and light and can wellinhabit sunny and hot bedrock, with temperature up to 50 °C. Higher fitness of C. acuminata in this habitat suggests that there are particular traits associated with its thermotolerance. Secondary metabolites may be involved in resistance against heat shock. It is well known that C. acuminata is a terpenoid indole alkaloid (TIA) producing plant. Its products, camptothecin (CPT) and 10- hydroxycamptothecin (HCPT), have been intensively studied in recent years. They have received wide attentions from pharmacologists and plant biologists all around the world due to their anti-tumor activities, which stems from their ability to inhibit DNA Topoisomerase I (Wall et al, 1966; et ability to inhibit DNA Topoisomerase I (Wall et al, 1966; Wani et al, 1986). As secondary metabolites, CPT and HCPT may play a crucial role during biotic and abiotic stresses, which pose great impacts on alkaloid biosynthesis and accumulations. The increase in alkaloid accumulation during seedlings development was observed and this shows that CPT and HCPT may play a defensive function for the plants during this vulnerable stage of their life cycle (Lopez-Meyer and Nessler, 1997). Heavy shading imposed on C. acuminata seedlings caused a shift of CPT accumulation from roots to buds. Subjecting plants to drought conditions would lead to increased biosynthetic activities in the alkaloid CPT (Liu, 2000). The current study was conducted to give a picture of the relationship between heat shock and alkaloid changes and the different responses of CPT and HCPT to heat shock in C. acuminata seedlings. This may provide some evidence that CPT and HCPT are differentially involved in the resistance of C. acuminata against external stress and stand for two unlike defensive strategies. 1 MATERIALS AND METHODS Camptotheca acuminata Decaisne seedlings used in the manipulation of heat-shock were three-month old constantly cultured in the highly automatic greenhouse of Northeast Forestry University in Harbin of China under the following conditions: temperature 25 °C, humidity 65%, and a light regime of 16 h. The average height of seedlings was 50 cm and the number of leaves per plant was about eight. The acquisition of controlled elevated temperature of plant was by putting soil-free C. acuminata seedlings into water-controlling container with constant temperature. There were two parameters, heat-shock time and temperature, to be considered during heat shock. We choose 2 h as the tentative time and adjusted the temperature within 34-46 °C at 2 °C intervals. According to the elementary experiments showing that peak values of CPT and HCPT contents appeared at about 40 °C, we fixed heat-shock temperature at 40 °C and adjusted time period from 1 h to 6 h at one-hour intervals for the following heat-shock trials. The upper three leaves were harvested and separated into three parts immediately after each manipulation, respectively for alkaloid, chlorophyll and malondialdehyde (MDA) level determinations. Parts for alkaloid measurement were oven-dried at 70 °C for 24 h, and those for chlorophyll and MDA determinations were made immediately after harvest as index of seedlings damaged extent. To investigate different responses of different organs to heat shock in alkaloid concentrations, young leaves (the upper three leaves), old leaves (the bottom three leaves) and barks were separated for alkaloid measurement. The determinations of chlorophyll and MDA contents were done by using the method of Lichtenthaler (1987) and Hadley (Draper et al, 1993), respectively. The alkaloid extraction and analysis was according to Liu et al (1998). The oven-dried leaf powder (from 60 mg to 1 g) was dissolved in 10 mL absolute ethanol (analytical grade). The low-frequency ultrasonic technology (250 W, 40 kHz) was used to extract CPT and HCPT about one hour. The ethanol extract was then centrifuged at 15 000 r/min, 10 min, at 10 °C for a High-Performance Liquid Chromatography (HPLC, Jasco, VG, England) and this HPLC system was equipped with a Waters ODS C18 reverse-phase column (250 mm×4.6 mm) and a photodiode-array detector set up to scan at 266 nm in the previous 8 min for HCPT and at 254 nm in the rest 12 min for CPT. Sample injection volume was 10 mL according to the presumable alkaloid content. The identification and quantification of the two alkaloids was performed at the base of the retention time and absorbance spectra of external standard sample of CPT (Sigma Chemical Co., St. Louis. MO 63178, USA) and HCPT (Sigma Chemical Co., St. Louis. MO 63178, USA). 2 RESULTS Alkaloid, MDA and chlorophyll contents were measured in C. acuminata leaves after two-hour heat-shock at different temperature treatments (Figs.1, 2). Heat shock led to an distinct rise in HCPT amount before 42 °C. During this process, HCPT level constantly increased step by step, followed by an abrupt rise. The peak value of HCPT concentration appeared at 40 °C and reached 3 363.338 µg/g DW, which was 6-fold of that in leaves of natural condition. Thereafter, there was constant decrease in HCPT content with enhancement of heat-shock temperature. In comparison, CPT seemed less heat-inducible according to varying extent with heat stress although it was influenced by external heat shock. It is surprising that there was a slight reduction of CPT concentration when heat-shock temperature was fixed at 34 °C and 36 °C. Then, at 38 °C, heat-shock induced a pop-up of CPT level, up to 2 000.000 µg/g DW. After that, CPT gradually rebounded into the bottom with a similar alkaloid level to the origin. On the other hand, the peak of CPT accumulation came earlier than that of HCPT. Heat shock in the present study actually caused damage to C. acuminata seedlings, because both MDA and chlorophyll displayed dramatic changes during this course (Fig.2). Chlorophyll level showed the trend of reduction, though evident declination did not emerge until heat-shock temperature had gotten to 38 °C. MDA level changing pattern was different from that of chlorophyll and it was revealed to have a constant increase after temperature reached 38 °C. The turning points of MDA and chlorophyll changes emerged at 42 °C, which was later than that of alkaloid. With prolong of heat-shock time, HCPT content ascended in response to heat stress and attained its peak value of 3 129.591 µg/g DW after two-hour shock. After that, intense declination of HCPT took place and at last recovered to the initial level (Fig. 3). In spite of a slight decrease at the beginning of heat shock, subsequently, CPT increased and reached its climax, 2 619.345 µg/g DW, also after 2-h stress. Furthermore, the total amount of HCPT and CPT displayed a visible promoter when heat-shock time was extended to 2 h. With further manipulation, CPT concentration started to descend. The responses of different organs and tissues of C. acuminata seedlings to heat shock were investigated by determination of alkaloid changes (Figs. 4, 5, 6, 7). Under natural developmental control, HCPT and CPT were heterologously distributed and allocated among young leaves, old leaves, buds and seedling barks, but remarkably homeostatic when viewed from a whole-plant perspective. Under heat-shock condition, alkaloids changed sensitively in buds and young leaves. Their responses in old leaves and seedling barks were characterized by relatively slight variation. In addition, HCPT experienced a more positive fluctuation all the time than CPT. In young leaves, peak value of HCPT, 2 860.218 µg/g DW, was 5-fold of the low value, 580.984 µg/g DW. While the CPT accumulation peak value was only 1.5-fold of the low point. However, what is more important was that the CPT peak always appeared before the peak of HCPT. 3 DISCUSSION The majority of known alkaloids are constitutively expressed as secondary metabolites and are considered to be part of a constitutive defense system (Robers and Wink, 1998). One classical example is nicotine, which is a highly toxic alkaloid and one of the most broadly effective plant defense metabolites in a number of Nicotiana species (Baldwin et al, 1994; Baldwin, 1999; Baldwin, 2001). The induced biosynthesis and accumulation patterns of CPT and HCPT by heat shock demonstrated that they were involved in plant defense against heat shock. They were sensitively inducible during the incubation of heat stress. In control circumstance, CPT and HCPT were in stable and low levels and a promotion of external temperature up to 34 °C triggered a slight decrease of CPT and a tiny promotion of HCPT. When the temperature was constantly turned up, HCPT tended to ascend, at the same time CPT displayed the traits of decline. CPT and HCPT attained their peak values at 38 °C and 40 °C, respectively. When heat-shock temperature rose to 42 °C, the physiological balance of C. acuminata was considerably disturbed in that MDA and chlorophyll levels changed remarkably. Before that, CPT and HCPT, however, had been largely mobilized and promoted to work as a defensive weapon. This kind of strategy was observed in C. acuminata seedlings encountering other stress like drought and shade (Liu, 2000). But, the elaborate mechanism that CPT and HCPT play a crucial role under heat and other conditions remains unclear and needs to be addressed in the future. As defensive compound, nicotine, accumulates in tissues with a pattern that is consistent with predictions of optimal defense theory, which argues that defense metabolites are allocated preferentially to tissues with high fitness value and a high probability of attack. Young leaves, stems, and reproductive parts tend to have the highest concentrations, roots and old leaves, the lowest (Baldwin, 2001). The distributions of CPT and HCPT in naturally developed C. acuminata are also under rigid tissue control and young leaves, barks, fruits and seeds are the main deposit places of alkaloids. The pattern of alkaloid distributions indicated from the contrary standpoint that CPT and HCPT are common defensive strategy, in that these young and reproductive tissues are especially susceptive to external attack or stress. More interestingly, responses of alkaloids to heat shock are also characterized by this type of allocation and accumulation. In a word, plants have evolved to mobilize and allocate limited source, alkaloids, to young and reproductive tissues, in order to protect themselves more effectively and survive from stress conditions. This is one of natural cost-benefit paradigm, making use of least alkaloid source (cost) to achieve high fitness (benefit). It has been demonstrated that nicotine, which is known to be firstly synthesized in roots and transported to buds and young leaves, is synthesized de novo from (15N)-nitrate in response to wounding of leaves (Baldwin et al, 1994). In Catharanthus roseus seedlings, catharanthine and advanced precursors of vindoline biosynthesis, such as tabersonine, may be synthesized in roots to be transported to leaves and stems, for elaboration into dimeric alkaloids and vindoline (Endo et al, 1987). It remains unclear if CPT and HCPT were biosynthesized in roots and transported to buds and leaves during development and stress conditions. In the present study, HCPT was more sensitive than CPT in response to heat shock. This shows that their patterns controlled by heat shock are different and HCPT biosynthesis is under more rigid environment-specific control than CPT. Different accumulation properties of CPT and HCPT during heat shock suggest that HCPT may be the direct defensive strategy for C. acuminata. CPT, however, is a universal tactic as nitrogen storage sink. The higher activity of HCPT, the hydroxyl derivative of CPT, is assumed to be associated with its hydrophilic property, which confers the better water-solubility, consequently, easier to be transported after being biosynthesized in specific organs and tissues. This is consistent with the discovery that most of the defense and signal compounds found in the vacuole are hydrophilic, polar, because they have to pass the tonoplast first in order to be accumulated in the vacuole (Robers and Wink, 1998). ACKNOWLEDGEMENTS The authors sincerely thank Prof. Gunter SWCHARZE, the visiting Professor of Northeast Forestry University from Germany, for his reviews and correction advice. REFERENCES
(Managing editor: WANG Wei) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03009f7.jpg] [as03009f5.jpg] [as03009f3.jpg] [as03009f1.jpg] [as03009f6.jpg] [as03009f4.jpg] [as03009f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}