 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
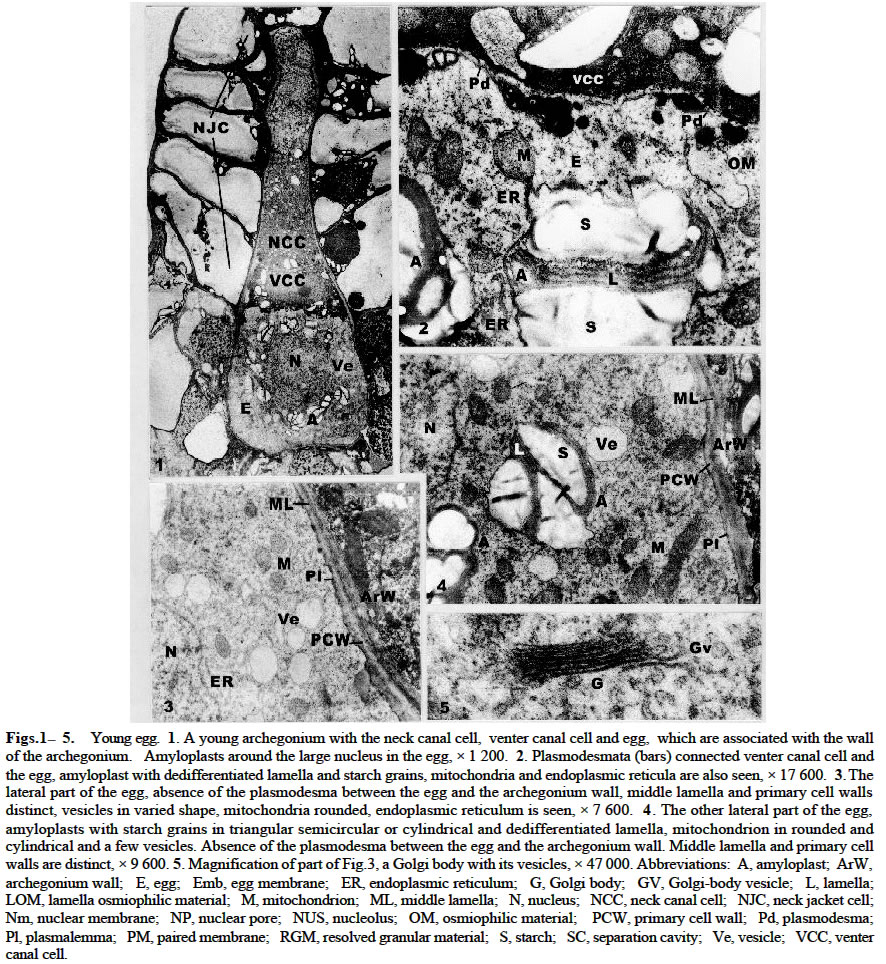
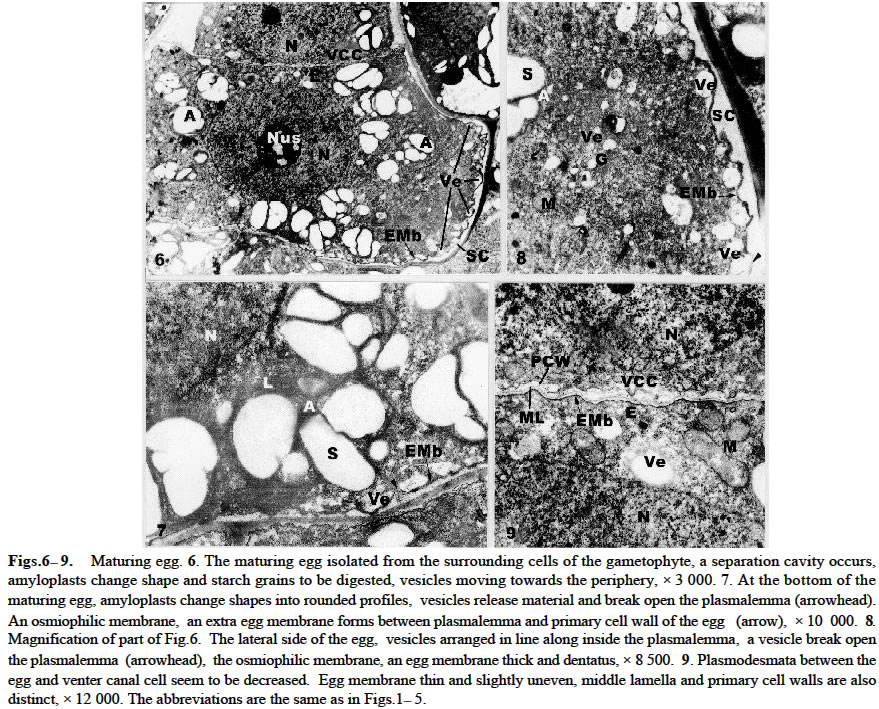
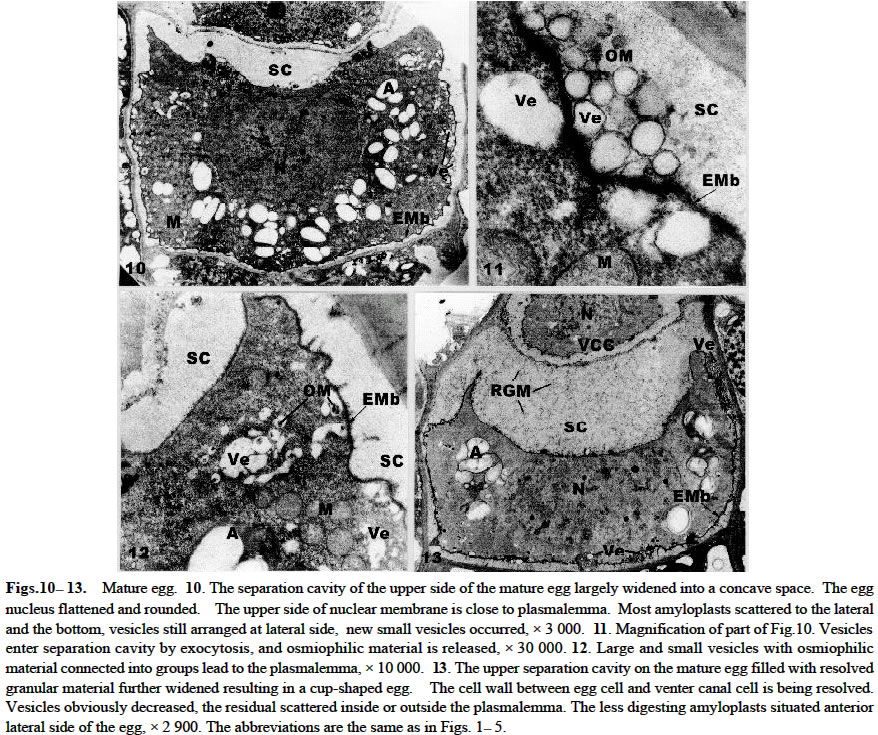
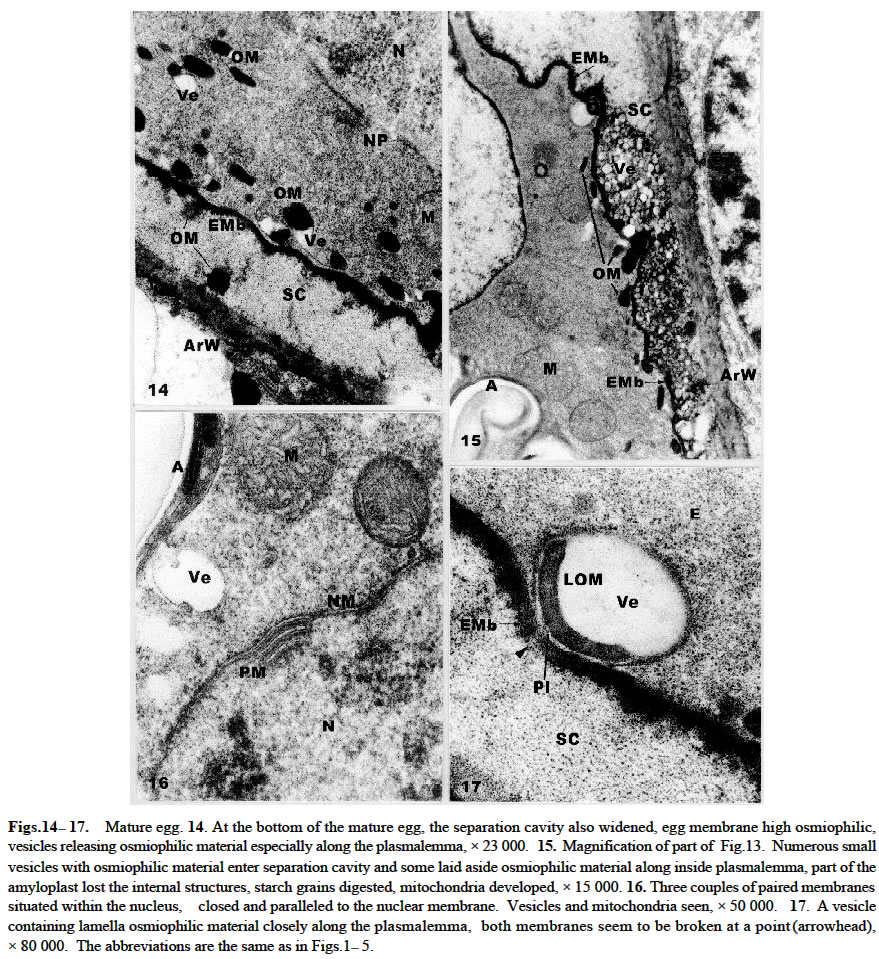
Acta Botanica Sinica 2003, Vol. 45, No. 7, pp.843-851 Ultrastructure of Oogenesis in Osmunda cinnamomea var. asiatica BAO Wen-Mei1, 3*, CAO Jian-Guo1, 2, DAI Shao-Jun1 (1. College of Life and Environmental Sciences, Harbin Normal University, Harbin 150080, China; Received: 2002-03-06 Accepted: 2003-03-22 Code Number: as03012 ABSTRACT The ultrastructure of oogenesis in the fern Osmunda cinnamomea L. var. asiatica Fernald has been studied by electron microscopy. During oogenesis, numerous vesicles not only moved towards the periphery, but also were arranged in line along the inside of plasmalemma, and in addition aggregated outside the plasmalemma by exocytosis. They released or excreted osmiophilic material. It was observed that a few vesicles containinglamellar osmiophilic materialsituated closely along the plasmalemma, seemed to break open. Simultaneously, a separation cavity between egg cell andarchegonium wallformed.Its width was broader than the other advanced ferns reported previously, and an extra egg membrane occurred outside around the plasmalemma of the egg, its thickness being greater than in Pteridium and Dryopteris. Amyloplasts around the nucleus were filled withlarge triangular semicircular or subelliptical starch grains, but as the egg matured they progressively decreased. Nucleus was large and flattened, and paired membrane in two to three couples laid within the nucleus, close and parallel to the nuclear membrane. No nuclear evagination was observed. Mitochondria seemed to have been undeveloped, but finally recovered normally. Key words: ultrastructure; oogenesis; Osmunda cinnamomea var. asiatica ; fern The developments of female gametes in the archegoniate have been studied by electron microscopy for a few decades, e.g. studies on Sphaerocarpus donnellii (Diers, 1966), Mnium undulatum (Barbier, 1972) and Marchantia polymorpha (Zinsmeister and Carothers, 1974) in the Bryophytes and detailed studies on Pteridium aquilinum in the Pteridophytes (Bell and Mühlethaler, 1962a; 1962b; Bell, 1963; 1972; Bell and Duckett, 1976). Studies of oogenesis of P. aquilinum were better known than those in any other ferns. Studies published on the other ferns have included: nuclear sheets of the egg and the nuclear evagination with acyl transferases in Dryopteris filix-mas (Bell, 1974; Cave and Bell, 1975), the nucleocytoplasmic interaction of the egg in Histiopteris incisa (Bell, 1980), and tubular elements in plastids of the egg in Pteris ensiformis (Bell, 1982). According to the Ching system (Ching, 1978a; 1978b), these belong to Leptosporangiopsida, which are more or less advanced species. It was not until 1986 that Bell reported the features of egg cells of living ancient ferns using electron microscopy. Todea barbara was studied, and its micrographs were taken, which exhibited the features of egg cells of the primitive ferns. It was mentioned that the egg cell of Osmunda resembled that of Todea. But no electron micrographs and description of it were presented. Reviews on the reproductive biology of Pteridophytes were well done by Bell (1970; 1979), DeMaggio (1977), Sheffield and Bell (1987) and Raghavan (1989), but these included very few references on the primitive ferns. Oogenesis of the fern related to Protoleptosporangiopsida (Ching, 1978 a; 1978 b) has not been studied which is different from that of the ferns of Leptosporangiopsida. This paper presents an ultrastructural description of developmental oogenesis in Osmunda cinnamomea var. asiatica, a primitive fern, belonging to Protoleptosporangiopsida. The egg at various developmental stages ( young, maturing and mature ) is studied in detail. As reported by Bell (1984), the ultrastructure of the egg cell of fern, although variable, does show a relationship to the evolutionary status and taxonomic affinity. 1 MATERIALS AND METHODS The spores of Osmunda cinnamomea L. var. asiatica Fernald were collected and identified in the late April 2000 from sporophylls of a plant growing on the edge of a forest from Laoshan Mountain, in Shangzhi County, Heilongjiang Province, China. The culture of archegoniate gametophytes has been obtained as previously described (Bao et al, 1986) with a minor change: the fresh green spores were washed with sterilized distilled water and sown directly. The culture medium consisted of a modified Knop's solution, solidified with 1.5% agar, autoclaved for 15 min at 15 lbs atmospheric pressure. Culture dishes at (20 ± 2) °C, under fluorescent light (ca. 2 000 lx) with continuous illumination. After about two months, archegonia occurred on the lower surface of the gametophytes. Slices were cut about 1 mm width in long strips from the archegonia regions of the gametophyte, and then plunged into 5% glutaraldehyde in 0.1 mol/L phosphate buffer at room temperature for 6-12 h. The materials were subsequently washed with their buffer and fixed in osmic acid (2% aqueous solution) for 2 h or overnight. They were then washed with the buffer, dehydrated in a graded acetone series and embedded in Epon 812. Specimens were thin-sectioned with a diamond knife on an LKB-Nova ultramicrotome and mounted on grid. They were stained at 20 °C with uranyl acetate for 30 min (7% aqueous solution) and then with lead citrate for 20 min. All specimens were observed under the JEOL-2000 electron microscope. 2 RESULTS The egg is contained within the archegonium. The whole archegonium develops from a single superficial cell, usually just behind the apical notch of the lower surface of the heart-shaped gametophyte. This cell gives rise to a column of three cells, the upper of which goes to form the neck of the archegonium and the innermost cell its basal are jacket cells. The cell in the middle is the primary cell. The division of the primary cell is strikingly unequal. Most of the cytoplasm remains with the lower cell, the central cell, and the one with the smallest cytoplasmic compartment becomes the neck canal cell. The central cell division is also unequal, producing the largest high cytoplasmic compartment large egg cell occupying the base of the archegonium, and the small cell next to a venter canal cell. The neck jacket cells, which are 6-celled high, about 80-100 µm and 50-60 µm in diameter, emerge from the surface of the gametophyte, and the venter jacket cells are hidden from the gametophyte (Fig.1) 2.1 Young egg The periphery of the young egg is associated closely with the wall of adjacent cells. The egg cell is short and cylindrical, about 30 µm in diameter, and possesses a large round nucleus with less regular shape. Chromatin is more dispersed in the nucleus than in the cytoplasm and elec-tron-opaque material is great amount (Fig.1). Numerous plasmodesmata connect the venter canal cell to the egg cell (Figs.1, 2), as well as those of the venter canal cell to the neck canal cell (Fig.1). Plasmodesma are absent between the cells of the axial row and the archegonium wall (Figs. 3, 4). The conspicuous feature of the young egg cell is the presence of numerous amyloplasts arranged in a region around the nucleus (Fig.1). The amyloplasts are almost filled with starch grains often in triangular, semicircular or cylindrical forms and lamellae being dedifferentiated in it (Figs. 1, 2, 4). The vesicles adjacent to the region of amyloplast in numerous profiles are dispersed. Some of them reach 1.0-1.5 µm in diameter and are bounded by double membranes(Figs. 3, 4). Many mitochondria are round and club-shaped in section. Double membranes and cristae are recognizable. The electron-opaque material in the mitochondria are more than that in the cytoplasm (Figs. 2-4). Few endoplasmic reticula (Figs. 2, 3) and Golgi body appear in the cytoplasm (Fig.5). 2.2 Maturing egg The volume of the maturing egg increases at its widest, reaching 35 µm (Fig.6). At first sight it appears to be isolated from the surrounding cells of the gametophyte by a space between the egg and archegonium wall, referred to as the separation cavity. It differs from those present at an earlier stage. It is still in close contact with the egg and venter canal cell (Fig.6). Under the electron microscope, the middle lamella appears as darkly staining material flanked on each side by primary cell walls ( Figs. 7, 9). The nucleus is still less regular as seen previously. There is no nuclear evagination appearing as reported in the ferns Pteridium (Bell, 1972) and Dryopteris (Bell, 1974; Cave and Bell, 1975). Many amyloplasts are scattered, but few are at the top of the egg (Fig.6). They become progressively dedifferentiated and lamellae are structureless; some of the starch grains seem to be digested (Figs. 6-8). A striking change at this stage is that there are small or large vesicles moving towards the periphery. Some of them contain osmiophilic material and seem to be discharged at the surface of the protoplast (Figs.7, 8). Simultaneously, a new osmiophilic membrane begins to form outside the plasmalemma of the egg, but not present in the adjacent cell (Figs. 6-8). This new extra egg membrane is probably analogous to that present around eggs of other ferns. The common wall between the egg and the cell of the gametophyte is well defined by the separation cavity and devoid of plasmodesma (Figs. 6, 7). After the formation of the egg membrane, the separation cavity expands rapidly and the maturing egg becomes largely isolated from the metabolism of the gametophyte. The egg membrane on the top of the egg is thin (Figs. 6, 9), but it is thick and dentatus around the lateral and lower surface of the egg (Figs. 6-8). The separation cavity and vesicles were tested for reaction with PAS. Both of them were stained in pink. The material within them, therefore may be a hydrated excretion by vesicles. Mitochondria, though, remain round and cylindrical with double membranes, and eral and the bottom, but not at the top of the cytoplasm. appear to have lost most of their internal structures (Fig.9). 2. 3 Mature egg In the first stage of the egg maturation, it is remarkable that the separation cavity of the upper side of the mature egg largely widens, forming a concave space wider than that of the other side of the egg (Fig.10). Nucleus is large and slightly irregular in outline. The upper membrane of the nuclear is very near to the plasmalemma of the egg. Amyloplasts decrease and are mostly scattered at the lateral and the bottom, but not at the top of the cytoplasm. Starches decrease and seem to have been digested (Fig. 10). Vesicles mostly containing osmiophilic material move towards the periphery. Some of them have entered into the separation cavity by exocytosis (Figs. 10, 11). In addition, a lot of vesicles, large or small, round or irregular, containing osmiophilic material are grouped and seem to lead to the plasmalemma. Mitochondria increase, most of which are round and well distributed (Fig.12). In the second stage, the egg completely matures. The separation cavity of the upper side of the egg, filled with resolved granular material, further widens to 8 µm, resulting in a cup-shaped egg cell (Fig.13). The separation cavity around the lateral side and the bottom of egg are also widened, the widest reaches 1.2 µm (Figs.13, 15). The cell wall between the egg and venter canal cell is being resolved, the venter canal cell seems to undergo a programmed cell death (Fig.13). Nucleus is more flattened than at the previous stage. Its transverse axis (ca. 17 µm) is longer than that of the longitudinal one (ca. 8 µm), occupying more than one third volume of the egg cell. The upper membrane of the nucleus is almost associated with the plasmalemma of the egg, and the bottom of the nucleus is closed to the plasmalemma of the egg cell (Fig.13). Paired membranes in two to three couples lie within nucleus close and parallel to the nuclear membrane (Fig.16). Amyloplasts decrease obviously and the residuals are all scattered in the anterior lateral area (Fig.13), lose most of the internal structures (Figs.13, 15). A few vesicles containing lamellar osmiophilic material distributed along the plasmalemma, both of them seem to be broken at a point on the opposite side (Fig.17). Numerous small vesicles are almost progressively disappearing from the cytoplast, but a lot of them were bounded with a double membrane and grouped in a large aggregates. They appear outside of plasmalemma in the separation cavity (Fig.15). In addition, there are many high osmiophilic materials which were probably excreted from vesicles, large or small, round or club-shaped inside the plasmalemma and amorphous outside the plasmalemma (Figs.14, 15) adding the egg membrane, in which the widest reaches 0.13 µm (Fig.17). Mitochondria are developed and recover the internal structures (Figs. 15, 16). 3 DISCUSSION The vesicles in the young egg often scatter near to the nucleus and mingle with the amyloplasts and mitochondria, and they are round or irregularly round, small or large. As egg matures, most of these vesicles move progressively towards the periphery and a new egg membrane forms. This process is similar in its essentials to those in other ferns such as Pteridium (Bell and Duckett, 1976) and Todea (Bell, 1986), but differs in detail from that in O. cinnamomea var. asiatica. The numerous vesicles not only move towards the periphery, but also are arranged in line along the inside of plasmalemma, especially at the lateral and the bottom of the egg. A few vesicles seem to break and open the plasmalemma (Figs.7, 8). As maturation proceeds, numerous new vesicles appear in the cytoplasm, containing osmiophilic material connected in a system extending from the cytoplasm outwards to towards the plasmalemma. Some of them are grouped in large aggregates by exocytosis and appear outside the plasmalemma (Fig.11). The osmiophilic materials, released and laid aside by vesicles, are often observed (Figs. 14, 15). Moreover, a few vesicles contain lamellate osmiophilic material inside the plasmalemma. Its membrane is in close contact with the plasmalemma. Both of them are broken at a point and probably it is releasing material outside the plasmalemma (Fig.17). The possibility is that the lamellate osmiophilic material through the break directly adding on the egg membrane. This process and its development in these vesicles have not been reported in the other ferns so far. It is suggested in the present study that the vesicles excrete or release the osmiophilic material inside or outside the plasmalemma adding and widening the egg membrane. It has been identified previously as consisting of lipid (Cave and Bell, 1974). The origin of these vesicles is obscure. Bell (1969) reported that the “opaque” vesicles in Pteridium were thought to be derived from the degenerated plastids and the “empty” vesicles from the degenerated mitochondria. There are no signs that the vesicles arise from the plastids and mitochondria in the present study. But numerous new small vesicles emerged from the cytoplasm, which were similar to Golgi vesicles in shape and size (Fig.5). They may originate from Golgi vesicles. In O. cinnamomea var. asiatica, the vesicles in the upper side of the plasmalemma are fewer than those at the lateral and the bottom of the plasmalemma. The egg membrane is thin and slightly uneven at the top of the egg, thick and dentatus at the lateral and lower surface of the egg. It is shown that the formation and width of egg membrane are closely related to the activity of vesicles. As to the thickening of the egg membrane around the egg, it is dissimilar to P. aquilinum in which it is rough at the top and smooth around surface of the egg (Bell and Duckett, 1976). Moreover, the egg membrane around the lateral side of the egg in O. cinnamomea var. asiatica is thicker (0.13 µm) than in P. aquilinum (0.1 µm at its widest side) (Bell and Duckett, 1976), but it may be close to T. barbara (0.15 µm) ( Bell, 1970). It is not strange with the relation to Osmunda and Todea, because both of them are primitive ferns. How to understand this phenomenon is still unknown. Although the egg membrane is thick and intact, it is fragmented at the point of entry of the spermatozoid ( Cave and Bell, 1974). Simultaneously, with the activity of the vesicles, a separation cavity occurs. When reaction with PAS was tested its product was colored pink. It closely resembles that of the vesicle material. This material may be a hydrated polysaccharide, which is also probably excreted from the vesicles. The separation cavity around the lateral side of the egg in O. cinnamomea var. asiatica is broader (the widest reaches 1.2 µm) than in Pteridium and Dryopteris (where its width does not exceed 0.5 µm) (Bell, 1970), but narrower than in T. barbara (2.5 µm in width) (Bell, 1986). The closer contact between the egg cell and the wall of the separation cavity may allow photosynthates to flow from the gametophyte into the fertilized egg and support its growth as the zygote germinates. The separation cavity being narrower, the soluble carbohydrates flow into the egg more rapidly. It seems that the width of the separation cavity of the egg in the above genera does show the narrower separation cavity representing the higher evolutionary status. In O. cinnamomea var. asiatica, the most conspicuous feature of the egg cell is in the presence of numerous large amyloplasts containing one or more starch grains around the nucleus (Fig.1). The starch grains often have triangular semicircular or subcylindrical forms with undeveloped lamellae (Fig.4). It is quite different from those of Pteridium (Bell, 1972) and Dryopteris (Bell, 1974; Cave and Bell, 1975), which are amoeboid proplastids containing little or no starch. In Dryopteris crassirhizoma (unpublished) it is found out that a few starch grains in the amyloplast are often club-shaped or round, and it is not numerous or conspicuous organelle in the egg. As the egg matures, starch grains are digested and separated, each forming into cylindrical or round grains inO. cinnamomea var. asiatica (Figs. 6, 10). In the fully mature egg, amyloplasts decrease (Fig. 13). Clearly, the growth and development of the egg is fueled by starch grains. The presence of numerous amyloplasts during its development is extraordinarily similar to the primitive ferns, such as T. barbara (Bell, 1986). It may be traced back to fern allies such as Equisetum which also manifests conspicuous amyloplasts in the egg, representative of a line of evolution extending from the Palaeozoic (Bell, 1986). It seems reasonable to consider the presence of numerous amyloplasts in the egg as a primitive feature retained in a number of ferns with a long history. However for the recent ferns, Pteridium and Dryopteris providing more energy-supplying carbohydrate is more advantageous than Osmunda's and Todea's starch grains. As each egg matures, the nucleus becomes flattened and its transverse axis elongates in O. cinnamomea var. asiatica. Within the nucleus paired membrane in two to three couples lie closed and parallel to the nuclear membrane (Fig.16). They were also seen in T. barbara (Bell, 1986), on the contrary, it has not been observed in Pteridium and Dryopteris (Bell, 1970). But its role in the developing egg is still unknown. Like Todea in the whole development process, no nuclear evaginations penetrating into the cytoplasm are observed, but this is a common occurrence in advanced ferns. The form of these evaginations depends upon the species, e.g. they are sac-like in P. aquilinum (Bell and Duckett , 1976), forming extensive sheets in D. filix-mas (Bell, 1974) and membrane systems in Histioptens incisa (Bell, 1980). Nuclear evaginations strengthen nucleocytoplasmic interactions in the developing egg preparing the egg cytoplasm to the embryo growth before fertilization. Without any evagination, it may put the primary ferns, Todea and Cinnamomea at a disadvantage, as the embryos grow. In P. aquilinum mitochondria are swollen and degenerated, the egg begins to form new mitochondria from detached nuclear evaginations (Bell and Mühlethaler, 1964; Bell and Duckett, 1976). In O. cinnamomea var. asiatica such degeneration of mitochondria has not been observed. They are only changed during the development and maturation of the egg unlike P. aquilinum. Since the egg is devoid of nuclear evaginations, mitochondria generated from the nuclear evagination are not observed in the present study. ACKNOWLEDGEMENTS Authors are grateful to Dr. James BRUSH for improvement of the manuscript. REFERENCES
(Managing editor: WANG Wei) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03012f6-9.jpg] [as03012f14-17.jpg] [as03012f10-13.jpg] [as03012f1-5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}