 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 864-870 Cloning and Analysis of a Disease Resistance Gene Homolog from Soybean WANG Bang-Jun1,2, ZHANG Zhi-Gang1, LI Xue-Gang2, WANG Yong-Jun1, HE Chao-Ying1, ZHANG Jin-Song1*, CHEN Shou-Yi 1* (1. Plant Biotechnology Laboratory, Institute of Genetics and Developmental Biology, The Chinese Academy of Sciences, Beijing 100101, China; 2. Central Laboratory, Southwest Agricultural University, Chongqing 400716, China) Supported by the State Key Basic Research and Development Plan of China (G1998010209). Received: 2002-05-20 Accepted: 2002-09-03 Code Number: as03014 Abstract: Conserved domains e.g. nucleotide binding site (NBS) were found in several cloned plant disease resistance genes. Based on the NBS domain, resistance gene analogs (RGAs) have been isolated previously and were used as probes to screen a soybean (Glycine max L. Merr.) cDNA library. A full-length cDNA, KR3, was obtained by screening the library and rapid amplification of cDNA ends (RACE) method. Sequence analysis revealed that the cDNA is 2 353 bp in length and the open reading frame (ORF) codes for a polypeptide of 636 amino acids with a Toll-Interleukin-1 receptor (TIR) and a NBS domain. Sequence alignment showed that it was similar to N gene of tobacco. The phylogenetic tree analysis of R proteins with NBS from higher plants was performed. The KR3 gene has low copies in soybean genome and its expression was induced by exogenous salicylic acid (SA). Key words: disease resistance gene homolog; nucleotide binding site; Toll-Interleukin-1 receptor; soybean
Plants have evolved elegant and complex systems to defend themselves against the invading of microbial pathogens, nematodes and insects. One of the effective defenses is characterized by a gene-for-gene interaction that requires a special plant resistance (R) gene and a corresponding pathogen avirulence (Avr) gene (Flor, 1971). More than 30 R genes have been characterized from different plant species that provide resistance to a variety of different pathogen and pest species by map-based cloning or transposon-tagging strategies (Hulbert et al, 2001). Five main classes of R genes have been defined according to the structural characteristics of their predicted protein products (Dangl and Jones, 2001). The majority of functionally described R genes are the nucleotide binding site-leucine rich repeat (NBS-LRR) type. The NBS domains are characteristic of various proteins with ATP/GTP binding activity and comprise the P-loop, kinase 2a, kinase 3a and GLPL motifs (Traut, 1994), while LRR domains play roles in the interaction of protein-protein (Kobe and Deisenhofer, 1994). Conserved motifs in NBS domain have been used to isolate resistance gene analogs (RGAs) from soybean (Kanazin et al, 1996; Yu et al, 1996; He et al, 2001; Penuela et al, 2002), common bean (Rivkin et al, 1999; Creusot et al, 1999), Arabidopsis thaliana (Aarts et al, 1998; Speulman et al, 1998), potato (Leister et al, 1996), lettuce (Meyers et al, 1998), maize (Collins et al, 1998) and cereals (Leister et al, 1999). These RGAs are useful in physical mapping and as gene candidates in positional cloning. In the present study, RGAs of soybean (He et al, 2001) were used as probes to isolate candidate R genes from Kefeng 1, an SMV and nematodes resistant soybean cultivar. 1 MATERIALS AND METHODS 1.1 Plant growth condition and nucleic acid isolation A soybean (Glycine max L. Merr.) cultivar, Kefeng 1, was grown in the
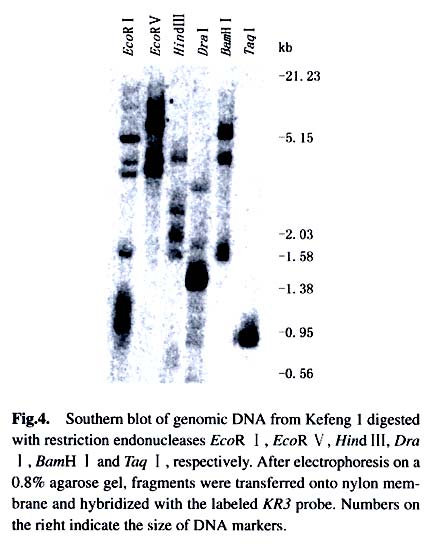
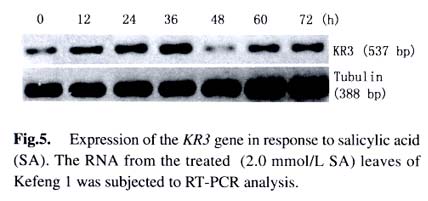
greenhouse under natural light. Leaves of 1.2 Library screening A lambda cDNA library was constructed with ZAPcDNA ® Gigapack® III Gold Cloning Kit according to the instruction manual (Stratagene). The mRNA was isolated from two-week-old leaves of Kefeng 1 after treating with SA for 24 h, 36 h, 48 h and 72 h, respectively. Thousands of recombinant phages of the library were plated on 150 mm plates and plaque lifts were carried out according to the manufacturer’s protocols. The phage DNA was cross-linked to the membranes under the UV light for 3 min. Hybridization was performed for 16 h at 65 °C with α- 32P-dCTP labeled probes in hybridization solution. Hybridization probe was prepared from soybean resistance gene analogs obtained previously (He et al, 2001). The filters were washed in 2 × SSC, 0.1% SDS for 20 min and then further washed in 1 × SSC, 0.1% SDS for 10-20 min at 65 °C. The positive plaques were cored from primary agar plates and screened for the second and third round to identify positive phages. Then the recombinant phages were excised and converted to plasmid for further analysis. The cDNA sequencing was carried out using the ABI 377 Automated Sequencer by Bioasia Biotech Co. 1.3 5'-RACE-PCR5'-RACE-PCR was performed because the isolated cDNA (KR3) was partial in its 5'-end. Two specific primers were designed according to the partial cDNA sequence of KR3 (GSP, 5'- CCAGCCAATTCTGCTACTTTCTGCAG-3'; NGSP, 5'-CTC GCGGTAGCAGCCAAGGATGGT-3'). The 5'-RACE-PCR was carried out according to SMART TM RACE cDNA Amplification Kit User Manual (Clontech). PCR reaction was performed for five cycles of 94 °C for 5 s, 72 °C for 2 min; five cycles of 94 °C for 5 s, 70 °C for 10 s, 72 °Cfor 2 min; 30 cycles of 94 °C for 5 s, 68 °C for 10 s, 72 °C for 2 min; and finally 72 °C for 10 min. Subsequently, the dilution of the PCR product was used in a nested PCR reaction. The nested PCR conditions were identical to the initial reaction. The products were analyzed on an 1.2% agarose/EtBr gel and the corresponding DNA band was recovered and directly sequenced by TaKaRa Biotech Co. 1.4 Southern blotting analysisSouthern blotting analysis was carried out as previously described (Chen et al, 1991). DNA samples prepared from Kefeng 1 were completely digested with restriction endonucleases EcoR I, EcoR V, Hind I, Dra I, BamH I and Taq I, respectively. Hybridization was performed for 16 h at 65 °C with α-32P-dCTP labeled KR3 cDNA as a probe. The filters were washed with 2×SSC, 0.1% SDS; 1×SSC, 0.1% SDS and 0.5 × SSC, 0.1% SDS for 15 min at 65 °C, respectively. 1.5 Reverse transcriptase-PCR (RT-PCR)The first-strand of cDNA was synthesized with 4 µg total RNA in a 20 µL of reaction volume using cDNA synthesis Kit according to the manufacturer's instruction (Promega). Two µL of the cDNA mix was used as a template in a 50 µL PCR reaction volume. After 5 min of denature reaction at 94 °C, the amplification was performed at 94 °C for 30 s, 56 °C for 30 s, 72 °C for 2 min for 30 cycles, with an extension of 10 min at 72 °C. The specific primers were designed according to the 3' sequence of KR3. The sense primer was 5'-CATTGCATGACTTGATTGAG-3' and the antisense primer was 5'-GAAAACCAATTGACTCTGAA-C-3'. A soybean tubulin gene, amplified with the primers 5'-AACCTCCTCCTCATCGTACT-3' and 5'-GACAGCATCA-GCCATGTTCA-3', was used as a control in this experiment. 1.6 Data analysisThe structure of KR3 domain was analyzed with EMBL Database (http://smart.embl-heidelberg.de). Multiple-se-quence alignment was made with the Lasergene software from DNAstar (Madison, Wis.). An unrooted phylogenetic tree was constructed using the Clustal W program (Thompson et al, 1994) that calculates distances based on progressive multiple alignment and uses the neighbor-join-ing method (Saitou and Nei, 1987) for tree construction. 2 RESULTS 2.1 Isolation and structure analysis of R gene homolog KR3 To isolate R gene candidate cDNA sequences, RGAs specific probes were used to screen a cDNA library constructed with the mRNA from SA-treated Kefeng 1. A total of 54 putative positives were obtained after screening approximately 2.7 million plaques. Several identical cDNAs were detected after restrictive enzyme digestion and electrophoresis analysis. Sequence analysis of the clones with longer inserts suggested that they had conserved domains related to R genes, but were partial in 5' cDNA sequences (data not shown). 5'-RACE-PCR was then carried out for KR3 and a product of 447 bp which overlapped KR3 129 bp was obtained subsequently. Thus, the full-length sequence of KR3 was obtained. KR3 (AF502079) is 2 353 bp in length including a 41 bp poly-A tail, and encodes a protein of 636 amino acids (Fig. 1). Structural analysis revealed that it had conserved motifs of R genes such as TIR and NBS, but had no LRR domain. Amino acid sequence alignment between KR3 and two known plant disease-resistance genes, tobaccoN gene and flax L6 gene, showed that the motifs such as the P-loop (GG(V/I)GKTT), kinase-2a (LL(I/V)LDD) and hydrophobic domain (HD, GLPL) were present in all the R proteins (Fig. 2). Further multiple alignments of amino acid sequences of several R gene analogs from soybean (KR1, L20a and L33) and four known R genes (N, L6, RPS2 and Xa1) from other plants were performed for the construction of a neighbor-joining phylogenic tree (Fig.3). KR3 was clustered withKR1, L20a and L33 before clustered with N gene. KR3 protein was more similar to tobacco N protein than the other R gene products and shared 28.1% sequence identity with N protein. This suggests that they might arise from close ancestral genes before speciation. 2.2 Southern blotting analysis Southern blotting analysis was performed in order to characterize the genomic organization of KR3 (Fig.4). The genomic DNA was digested with EcoR I, EcoR V, Hind III, Dra I, BamH I, Taq I, respectively, and subjected to hybridization. Several hybridizing bands appeared on the blot. Considering the fact that there were two EcoR I sites, two Hind I sites, two Dra I sites, two BamH I sites, six Taq I sites and no EcoR V sites in KR3 gene, KR3 was probably in low-copy numbers in soybean genome. 2.3 Expression of KR3 in soybean leaves Due to the very low abundance of KR3 gene, RT-PCR was used to examine its expression in soybean in respond to SA treatment. Total RNA was treated with DNase I (GIBCOL BRL) to avoid the genomic DNA contamination before reverse-transcription. The expected 537 bp fragment of KR3 and 388 bp fragment of tubulin were obtained (Fig. 5). It can be seen that, under normal condition, KR3 was expressed at a relatively low level in soybean leaf. However, application of 2.0 mmol/L SA to the plants resulted in accumulating of KR3 mRNA. The mRNA level reached a peak at 36 h after initiation of the SA treatment and then tended to decline, with fluctuations. 3 DISCUSSION Study of R gene is one of the key subjects of plant science. Cloning and characterization of R genes can not only facilitate the understanding on mechanism of interaction between host and pathogen but also provide mulating of KR3 mRNA. The mRNA level reached a peak at opportunities for producing crop varieties with increased disease resistance. Identification of RGA sequence from cDNA library would illuminate the full genetic structure of RGAs and correlate RGA-expressed sequences with known R genes by genetic mapping and physical association. Several full-length R gene candidates have been isolated from soybean (Graham et al, 2000; Hayes et al, 2000). In the present study, KR3 belongs to the class of plant R genes that contain TIR and NBS domains. The TIR is involved in rapid signal transduction in mammals and Drosophila (Hashimoto et al, 1988; Sims et al, 1989). NBS domain is necessary for the function of ATP or GTP binding. KR3 closely resembles N gene that confers resistance to tobacco mosaic virus (TMV). Sequence identity of the two genes is 28.1%. Conservation of NBS homologs may reveal the pace of evolution of R gene homologs. LRR domain is assumed to be Avr protein recognition site and is common to most cloned plant R genes. But in the flax L6 and tobacco N resistance genes, transcripts of truncated genes containing no or little LRR region have been observed (Whitham et al, 1994; Ayliffe et al, 1999). Truncated versions of resistance gene analogs have been identified in soybean. These truncated products have been presumed to have important role in disease resistance (Graham et al, 2000). The deduced protein of KR3 gene has no LRR domain. Thus, it is possible that KR3 may also be a truncated transcript in resistance gene families. Alternatively, KR3 may be a new type of R gene. SA, a product of the phenylpropanoid pathway, is involved in the expression of localized hypersensitive reaction (HR) as well as systemic acquire resistance (SAR) (Raskin, 1992). Application of exogenous SA has long been known to activate expression of pathogenesis-related (PR) genes and to induce resistance to plant diseases (White , 1979; Ward et al, 1991). Exogenous applied SA was observed to induce the genes that were expressed relatively early in the inoculated leaves of TMV-resistant tobacco (Guo et al, 2000). In the present study, the expression of KR3 was also induced after the treatment of exogenous SA, suggesting that KR3 may be involved in SA-regulated defense responses. Although the NBS domain was present in most plant R proteins, similar structural features were also present in the proteases CED-4 from C. elegans and APAF-1 from human, which have been reported to involve in the programmed cell death (PCD) (Shirasu and Schulze-Lefert, 2000). HR in plants is similar to PCD in animals, thus these structural similarities may reflect to some extent the functional similarities between KR3 and the PCD component. REFERENCES
(Managing editors: BAI Yu-Hong, HE Ping) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03014t2.jpg] [as03014f1.jpg] [as03014f3.jpg] [as03014f2.jpg] [as03014t1.jpg] [as03014f5.jpg] [as03014f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}