 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
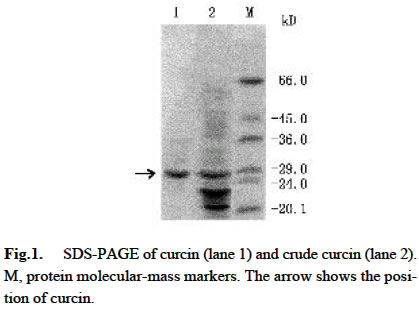
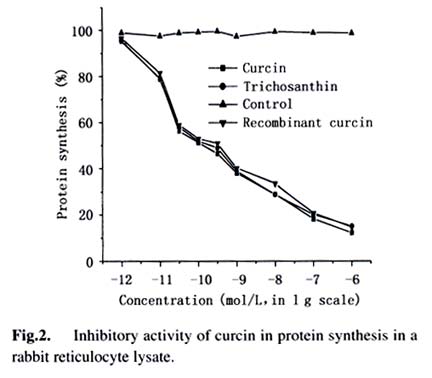
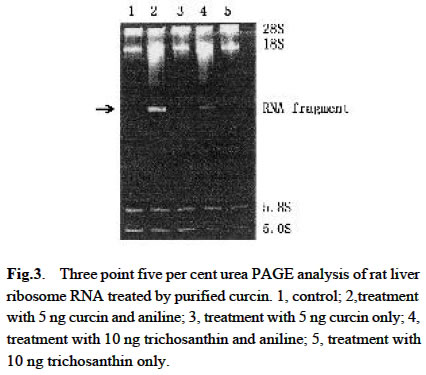
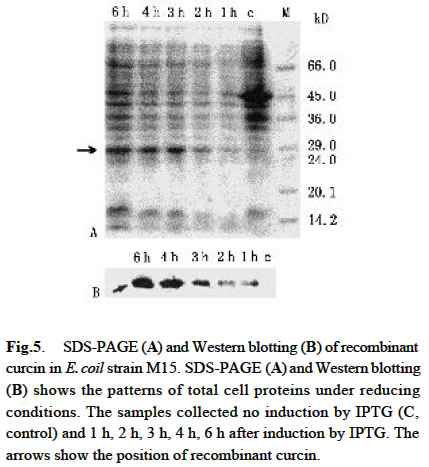
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 858-863 Cloning and Expression of Curcin, a Ribosome-Inactivating Protein from the Seeds of Jatropha curcas LIN Juan, CHEN Yu, XU Ying, YAN Fang, TANG Lin, CHEN Fang* (College of Life Sciences, Sichuan University, Sichuan 610064, China) Received: 2002-03-11 Accepted: 2002-08-26 Code Number: as03014 ABSTRACT Curcin, a ribosome-inactivating protein with a molecular weight of about 28.2 kD, which stronglyinhibits the protein synthesisin rabbit reticulocytelysate system with an IC50 value of about (0.19 ± 0.01) nmol/L, was purified from the seeds of Jatropha curcas L. The protein has the activity of rRNA N-glycosidase. Degenerate primers were designed based on the N-terminal partial sequence from purified curcin. The full-length curcin cDNA by RT-PCR and 5'-RACE was cloned. The deduced amino acids sequence indicates that a preprotein with 293 amino acid residues is first translated and then processed to a mature protein with 251 amino acids. The deduced amino acids sequence shares homology of 33% and 57% to those of type I ribosome-inactivating proteins (RIPs) and A chain of type II RIPs, respectively. The sequence encoding mature curcin was integrated into the pQE-30 vector for expression in Escherichia coil strain M15 (pREP4). The purified recombinant curcin was able to inhibit protein synthesis in rabbit reticulocyte lysate system. Key words: Jatropha curcas ; curcin; RNA N-glycosidase; cloning; in Escherichia coil expression Ribosome-inactivating proteins (RIPs) from plant are toxins that can inhibit protein synthesis in eukaryotic cells by catalytically damaging ribosomes (Barbieri et al, 1993). With rRNA N-glycosidase activity, when entering a cell the protein can enzymically cleave the specific glycosidic bond of an adenine residue (position 4 324 on the conserved GAGA loop of rat liver 28S rRNA), thus inhibiting protein synthesis by interfering with the binding of the 28S RNA to elongation factors (Peumnas et al, 2001). There are two types of RIPs. Type Ⅰ RIPs are a kind of single-polypeptide proteins with molecular masses of about 23-32 kD and a pI of 8-10, whereas type II RIPs (~60 kD) usually consist of two chains (A and B chain) connected by a disulphide bond. RIPs are believed to be involved in plant defense systems and have anti-fungal and anti-viral activities as well (Krawetz and Boston, 2000). In addition, RIPs are of great interest in theory and practice (Shaw et al, 1991). They have been used as a mid-term aborting agent (Cheung et al, 1989) and for treatment of hydatiform moles (Lu and Jin, 1990), and may have the potential in the development of future therapeutic medicines for AIDS (McGrath et al, 1990) and as anti-tumor agents (Wang et al, 1991). Here we describe the purification, characterization of curcin, a new RIPs from seeds of Jatropha curcas in Euphorbiaceae, molecular cloning and expression of its cDNA. This study is useful for revealing the relationship between structure and function of the RIPs and its use in medicine. 1 MATERIALS AND METHODS 1.1 Plant materials The immature seeds of Jatropha curcas L. were obtained from Panzhihua City, Sichuan Province, China, and immediately frozen in liquid nitrogen after removing seed pods and seed coats and keep at –70 °C until use. 1.2 Purification of curcin All purification procedures were performed at 4 °C. One hundred and twenty grams of the seeds of J. curcas was homogenized with a Waring blender by using 50 mmol/L sodium phosphate buffer (containing 0.2 mol/L NaCl, pH 7.2), and then extracted for 24 h. After centrifugation of homogenate at 10 000 r/min, solid (NH3)2 SO4 was added to the supernatant till to 60% saturation. After being left for overnight, the precipitates were collected by centrifugation at 12 000 r/min for 20 min, and then redissolved in 5 mmol/L sodium phosphate buffer (containing 0.2 mol/L NaCl, pH 7.2). Then the solution was dialyzed with the same buffer. At the end of dialysis, a brown precipitate was present in the bags, then removed out by centrifugation. The brownish supernatant referred to as crude curcin. The crude curcin was purified by CM-52 column and Sephadex G-100. Its purified fraction was analyzed by SDS-PAGE. 1.3 N-terminal amino acid sequence analysis For N-terminal amino acid sequencing, the protein in gels was transferred to polyvinylidene difluoride (PVDF) membrane by electroblotting, which was carried at a constant voltage of 100 V at 4 °C for 3 h. The blot was strained and individual bands were cut out, eluted from the membrane and subjected to N-terminal amino acid sequencing by using an ABI procise® 491 protein sequencer (Applied Biosystems). 1.4 Cell-free translation-inhibitory activity Protein synthesis was measured with a lysate of rabbit reticulocytes. Rabbit reticulocyte lysate was prepared as described by Sambrook et al (1989) and protein synthesis was assayed as Merrick (1983) and Sambrook et al (1989). The test sample (10 µL) (negative: trichosanthin purchased from Sigma Chemical Co., USA; positive: H2O) was added to 1 0 µLof hot mixture (reaction mixture contained, final volume of 1 mL: 250 µL 100 mmol/L phosphocreatine; 100 µL amino acids (Q, W, D, N, A, G 2 mmol/L, other 1 mmol/L, minus Leucine); 20 µL 100 mmol/ L GTP; 50 µL 10 mmol/L ATP; 200 µL 2 mol/L KCH3COO (pH 7.5); 10 µL 1 mol/L Mg (CH3COO) 2; 50 µL 10 g/L creatine phosphate kinase and 320 µL H2O), 25 µL working rabbit reticulocyte lysate containing 0.1 mmol/L hemin, 2 µL [3H] leucine (2 µCi) and 3 µL H2O. The reaction was performed at 37 °C for 30 min. One volume of the reaction mixture was then added to 40% trichloroacetic acid with 2% casein hydrolyzate in a 96-well plate to precipitate radioactively labeled protein. The precipitate was collected on a glass fiber (Whatman GF/A filter), washed and dried with absolute alcohol passing through a cell harvester attached to a vacuum pump. The filter was suspended in scintillant (4% POP-0.04% POPOP) and counted in an LS 5801 Backman liquid scintillation counter. 1.5 Assay for N-glycosidase activity The assay was performed in accordance with the procedure of Endo et al (1987), but with slight modifications. Rat liver ribosomes were prepared as described by Speeding (1990). The reaction mixture containing sample (curcin or trichosanthin) made up to 80 μ L with buffer A (20 mmol/L Tris-HCl (pH 7.6), 50 mmol/L KCl, 5 mmol/L MgCl2, 1.5 OD ribosomes) was incubated at 37 °C for 10 min. The reaction was stopped by addition SDS to a final concentration of 0.5%. The mixture was extracted with chloroform: isoamyl alcohol (24:1 (V/V)), and precipitated with cold absolute ethanol and 3 mol/L CH3COONa, and treated with acidic aniline (formed by mixing 1 volume aniline, 1 volume acetic acid and 3.5 volume DEPC-water). The reaction products were analyzed by using 3.5% urea-PA GE. The RNA bands were visualized on a UV transilluminator and photographed with bio imaging systems (Syngene, a division of Synoptics Ltd.). 1.6 Sequence analysis Total RNA was prepared from the seeds, which were ground in liquid nitrogen, and then extracted with the Plant RNA Mini Kit (Watson). Based on N-termined amino acid sequence, cDNA 3' end was obtained with the One Step RNA PCR Kit (AMV)(TaKaRa). Based on cDNA 3' end sequence, cDNA 5' end was obtained using the SMARTTM RACE cDNA Amplification Kit (Clontech). Subcloning and sequencing of PCR products were performed by using standard protocols (Ye et al, 2001). 1.7 Construction of expression vector The coding region of curcin cDNA was cloned into the bacterial expression vector pQE-30 with a pair of specific primers, JP1 (5'-AACGCATGCGCTGGTTCCACTCCA ACTTT-3') and JP2 (5'-ATACGTCAGATACATTGGAAAG ATGAGGA-3'), which had Sph I site and Pst I site respectively. The cDNA encoding curcin was obtained by using PCR amplification in the same manner as that for the cDNA 3' end. The PCR products and vector pQE-30 were digested by Sph I /Pst I. Digested products were gel-purified using Gel Purification Kit (Watson). Ligation, transformation and screening were carried out by standard protocols. 1.8 Expression and purification of the curcin in Escherichia coli A single colony of E. coli stain M15 (pREP4) harboring the expression plasmid pQE-30 was inoculated into 50 mL of LB medium containing ampicillin (final concentration 100 mg/L) and kanamycin (final concentration 25 mg/L), shaking incubated at 37 °C until an OD600 value of 0.6-0.7. The cells were collected by centrifugation and resuspended in 50 mL of fresh LB medium and inoculated into 500 mL of LB medium. The incubation was carried out under the same condition as described above. When the OD600 value reached 0.6, IPTG was added to a final concentration of 1 mmol/L. The culture was continued for 6 h. The expression level of the protein was assessed by analysis of total protein on 12% SDS-PAGE gel followed by Coommassie Brilliant Blue R250 staining. The recombinant curcin was purified by using Ni-NTA agarose affinity chromatography according to the manual of QIA expressionist (Qiagen). 1.9 Western blotting analysis Western blotting was conducted using RGS-His primary antibody and rabbit anti-mouse secondary antibody conjugated with alkaline phosphatase (AP). 2 RESULTS 2.1 Purification of curcin Curcin was purified to homogeneity from seeds of J. curcas by two simple steps and its molecular mass is about 28.2 kD by SDS-PAGE (Fig. 1). 2.2 Inhibitory activity of curcin As shown in Fig.2, the purified curcin and trichosanthin strongly inhibited protein synthesis in rabbit reticulocyte lysate, and their values of IC50 are (0.19±0.01) nmol/L and (0.22 ± 0.02) nmol/L, respectively. The value of trichosanthin IC50 corresponds to previous findings (Barbieri et al, 1993). 2.3 N-glycosidase activation The N-glycosidase activity of curcin was examined by incubating ribosome with various amounts of curcin or trichosanthin, and the extracted rRNA was analyzed by PAGE (Fig.3). As shown in Fig. 3, curcin clearly exhibited rRNA N-glycosidase activity. At 5 ng curcin only, the protein produced a diagnostic RNA fragment of approx. Four hundred and fifty nt (lane 2), similar to that found in 10 ng trichosanthin (lane 4). Treatment without curcin or trichosanthin (lane 1) or without anline (lane 3, 5) had no effect on rRNA as compared with that treated with curcin or trichosanthin and aniline. The arrow indicates the position of the RNA fragment. These results suggest that curcin has a RNA N-glycosidase activity as trichosanthin does. Curcin as an active RNA N-glycosidase could strongly inhibit protein synthesis in a cell-free translation system. 2.4 N-terminal amino acid sequence of curcin Thirty-two amino acids at N-terminal were sequenced as follows: A-G/Y-S/K-T/A-P/D- T-L-T-I-T-Y-D-A-T/A-A-D-K-K-N-Y-A-Q-F-I-K-D-L-R-E-A-F/A-G. Among these amino acids, some are uncertain. Comparison of the 15A 30A with other RIPs reveals that there are similarities among them. Therefore, the primers were synthesized according to underlined amino acids deduced from nucleotides. 2.5 Full-length curcin cDNA sequence Full-length cDNA and its deduced amino acid sequence were obtained by analysis of cDNA 3' end and cDNA 5' end sequence (GenBank AY069946). The deduced amino acid sequence indicated that an immature protein with 293 amino acid residues was firstly translated and then processed to a mature protein with 251 amino acids. The gene could encode a protein of 27.76 kD, which closes to that observed on SDS-PAGE of purified curcin protein. Two Nglycosylation sites are at Asn-266 and Asn-274, and the following amino acids are Asn-Try-Thr and Asn-Val-Thr. Unlike other RIPs, such as trichosanthin and momorcharin, curcin contained one cysteine residue, located at Cys-209. 2.6 Sequence similarity and comparison among various RIPs From the sequence information available, it is clear that the type I RIPs and A chain of type II RIPs are homologous and evolutionarily related. Amino acid residues of signal peptide are less conserved, but seven amino acid residues are conserved among curcin, ricin A chain, and gelonin from Euphoribaceae (Fig. 4). However, there was no significant homology between RIPs from Euphoribaceae and Cucurbitaceae. Based on the results, we infer that amino acids of N-glycosidase active site are conserved in different species, but amino acid residues of single peptide are different among families. It is interesting to note that all amino acid residues forming the active site of the trichosanthin are conserved in curcin (Tyr-20, Phe-23, Arg28, Tyr-76, Tyr-117, Gly-134, Ala-155, Glu-167, Arg-170, Asn-197, Trp-199, Leu-244), with the exception of one residue, i.e. Gln-156 of trichosanthin is replaced by Glu-163 of curcin (Fig. 4). The Gln and Glu are similar in property except that Gln has an extra — NH3 in side chain. These residues are consistent with the RIP activity, which might constitute the active site region of curcin. 2.7 Expression of curcin in E. coil SDS-PAGE patterns of total cell protein visualized by Coommassie Brilliant Blue R250 Staining showed the time course of the target protein's expression (Fig. 5 A). Its maximal level was achieved about 3 h after IPTG induction. The expressed protein had an apparent molecular mass of about 28 kD by SDS-PAGE, corresponding to the cDNA-deduced amino acid sequence of the mature type of curcin. Western blotting analysis showed that the induced samples had positive signal at 28 kD position, where there is no signal in uninduced sample. This result further confirmed that the induced E. coil had expressed recombinant protein with 6 × His-tag (Fig.5B). 2.8 Cell-free translation-inhibitory activity of recombinant curcin As most of the expressed proteins, the recombinant curcin was in the soluble fraction. It was purified under native conditions by a single Ni-NTA affinity column. After the elution, it could be purified to about 90% purity. Like native curcin, the recombinant one strongly inhibited protein synthesis in rabbit reticulocyte lysate, and the value of IC50 was (0.32 ± 0.04) nmol/L (Fig. 2). 3 DISCUSSION J. curcas is widespread in tropical and subtropical areas, planted as “living fences” around fields and settlements. It can be easily grown on almost any soil and therefore prevents the soil from erosion. It is abundant in southwest and southeast of China. Most parts of the plant and their extracts are used in traditional medicine for the antimicrobial properties (Thomas, 1989). Extracted oil shows insecticidal effects against crop pests such as Helicoverpa arimgera and Sitophilus zeamays (Solsoloy, 1995). The toxicity of the whole seed form J. curcas has been known for a long time. Felke (1914) isolated a toxic protein from the seeds of J. curcas and designated as “curcin”. He proposed that curcin was a kind of toxalbumin. Wehmer (1931) and Schwarting (1963) considered that the curcin was associated with ricin and abrin respectively. Stripe et al (1976) observed that the curcin was toxic to animals and inhibited protein synthesis in a cell-free system, although it was 1 000 times less toxic than ricin and abrin, but there was no toxins in whole cell. In our study, we not only present the molecular cloning of curcin cDNA, but also report its protein purification and inhibitory protein synthesis activity in a cell-free translation system. Thus, the curcin can be used as an important tool in biological and medical fields. Curcin contains one cysteine residue, it maybe directly form a disulphide bond with an activated antibody. A disulphide linkage is usually thought to be essential for maximal cytotoxicity. Most of type I RIPs have not any free cysteine residue. Therefore, the curcin is a novel protein, and RIP of containing cysteine, which might be ideal for the preparation of immunoconjugates with great potential as a chemotherapeutic agent for the treatment of various cancers or AIDS. This result shows that the curcin has similar molecular modeling on their structure as compared with the other RIPs. We deduce that the biological activity of the curcin might be the same as the other RIPs. Anti-tumor activity of the curcin has been found. Its potential application in biological, agricultural and medical fields will be found in the future. REFERENCES
(Managing editor: HAN Ya-Qin) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03015f2.jpg] [as03015f1.jpg] [as03015f4.jpg] [as03015f3.jpg] [as03015f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}