 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Acta Botanica Sinica, Vol. 45, No. 7, 2003, pp. 832-842 Ultrastructure of the Blepharoplast and the Multilayered Structure in Spermatogenesis in Osmunda cinnamomea var. asiatica CAO Jian-Guo1, 2, BAO Wen-Mei1, 3*, DAI Shao-Jun1 (1. College of Life and Environmental Sciences, Harbin Normal University,
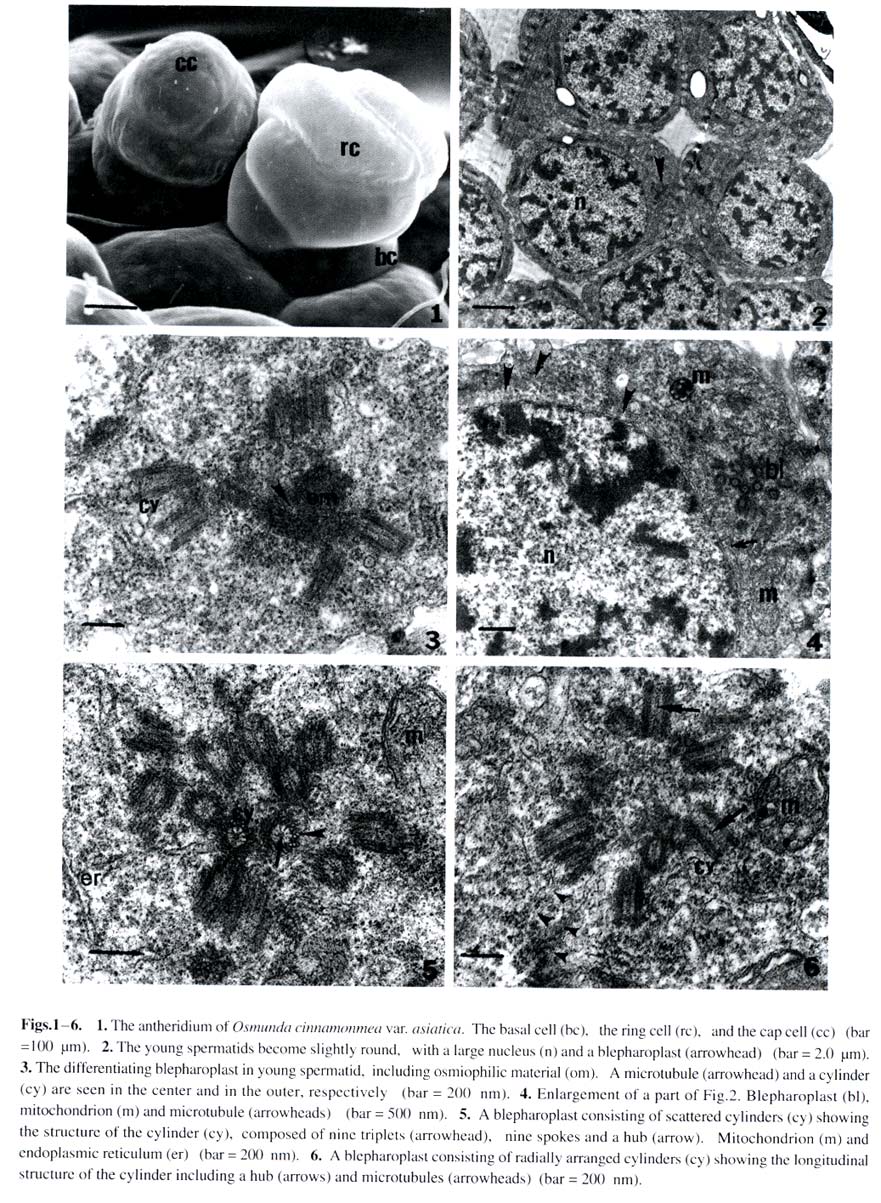
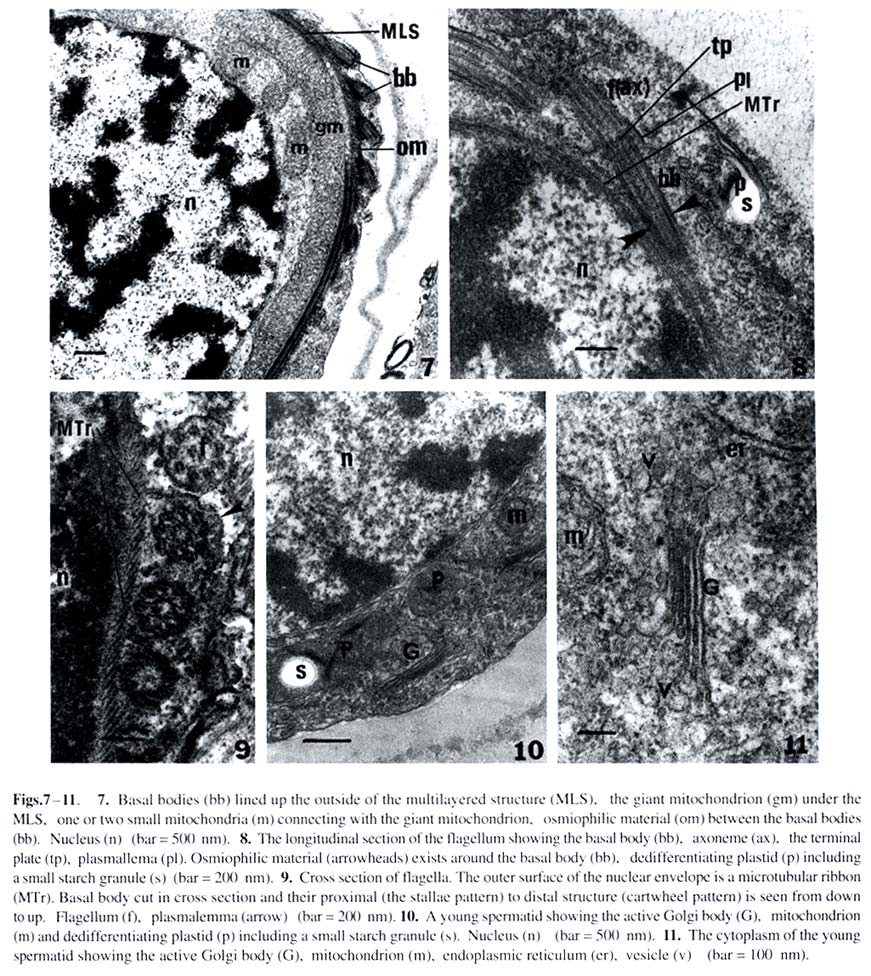
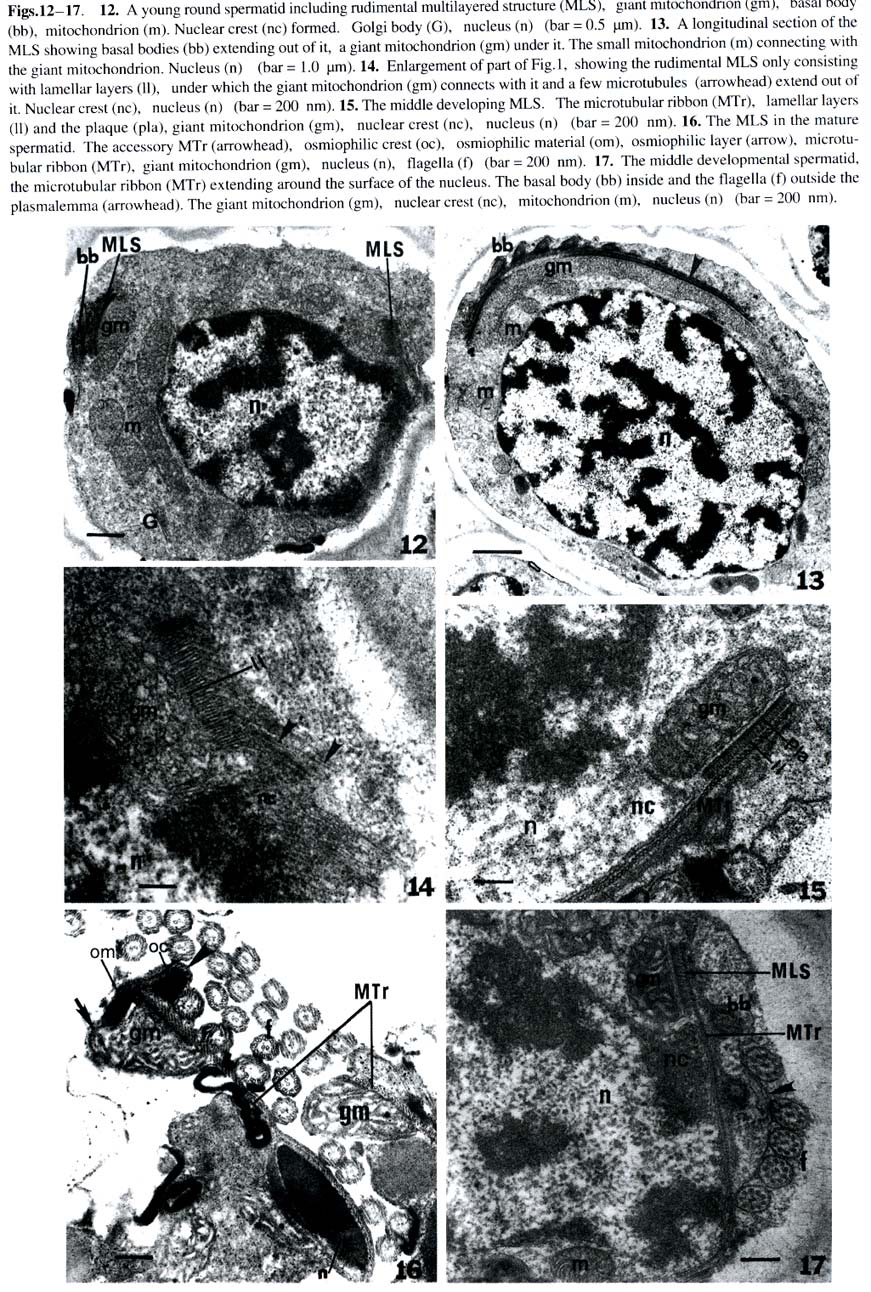
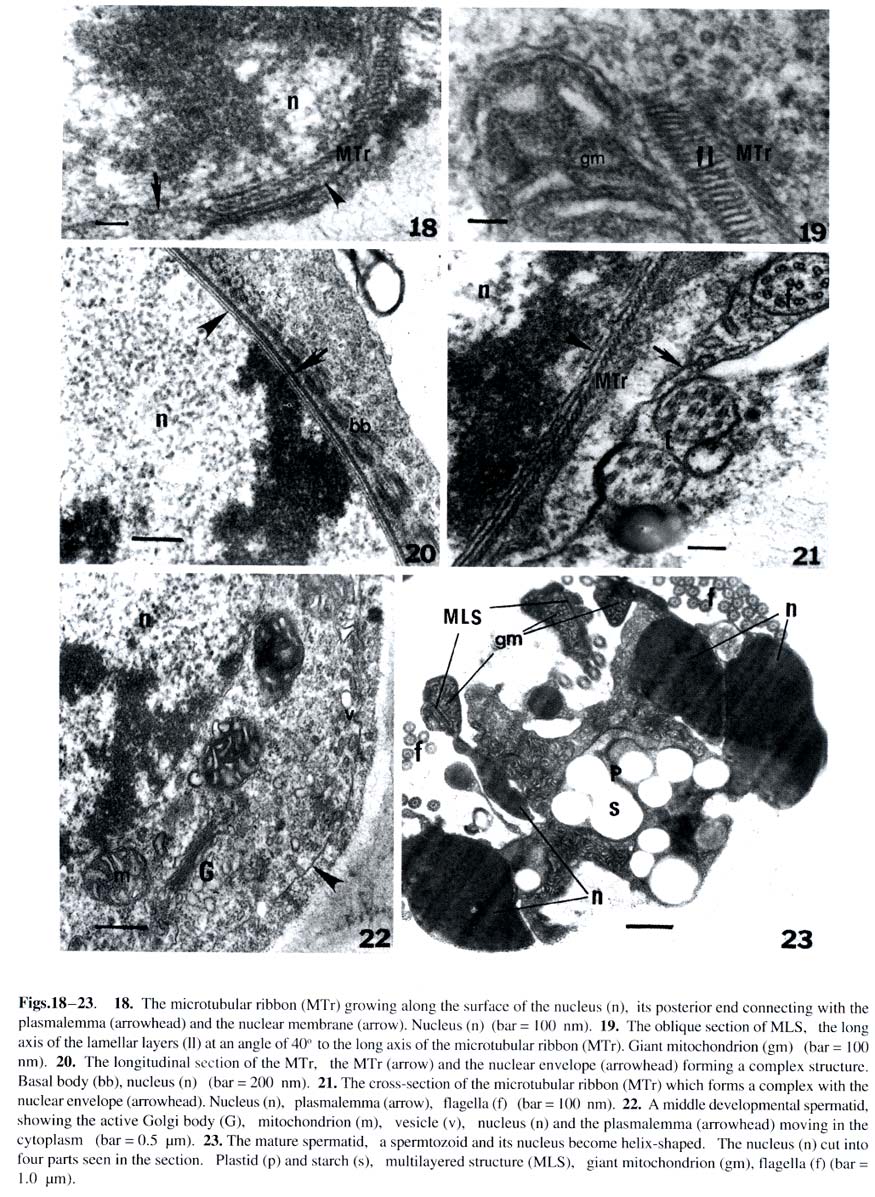
Harbin 150080, China; Received: 2002-02-25 Accepted: 2003-03-20 Supported by the National Natural Science Foundation of China (39774213) and the Heilongjiang Provincial Natural Science Foundation. Code Number: as03019 Abstract: The ultrastructure of the blepharoplast and the multilayered structure (MLS) in the fern Osmunda cinnamomea var. asiatica Fernald have been studied by electron microscopy with respect to spermatogenesis. The blepharoplast appears in the young spermatid. The differentiating blepharoplast is approximately a spherical body, which is composed of densely stained granular material in the center and some cylinders outside of it. The differentiated blepharoplast is also a sphere, but without the densely stained material in the center, consisting of scattered or radially arranged cylinders. The MLS seen in the spermatid lies between the basal bodies and the giant mitochondrion. In the early developmental stage, the MLS only consists of lamellar layers, each of which runs parallel to one another and forms a strip. In the mid stage, the MLSis composed of the microtubular ribbon (MTr), thelamellarlayers and alayer of plaque. In the late stage, the MLS forms accessory band, osmiophilic crest and a layer of osmiophilic material. The MTr grows out from the MLS and extends along the surface of the nucleus to unite with the nuclear envelope in a complex. The basal body coming from the cylinder produces the axoneme of the flagella in the distal end and the wedge-shaped structure in the proximal end, respectively. In the present study, the ultrastructural features of blepharoplast andthe MLS of the protoleptosporangiopsida fern, O. cinnamomea var. asiatica , have been described and compared with those of other kinds of pteridophytes in detail. The lamellar layers appearing before the formation of the MTr was found and reported for the first time. Key words: Osmunda cinnamomea var. asiatica ; blepharoplast; multilayered structure; ultrastructure The blepharoplast and the multilayered structure (MLS) are two organelles, which arise de novo in the spermatogenesis of the archegoniate plants. They are important in the formation and development of the spermatozoid. Doonan et al (1986) used anti-tubulin antibodies to study the blepharoplast in the spermatogenesis of Platyzoma microphyllum, indicating that the blepharoplast is the microtubule- organizing center (MTOC). In recent 20 years, the blepharoplast during spermatogenesis of Coleochaete scutata (Graham and Mcbride, 1979), Marchantia polymorpha (Carothers and Kreitner, 1968), Aulacomnium palustre (Bernhard and Renzaglia, 1995), Lycopodium cernuum (Robbins and Carothers, 1978), Selaginella australiensis (Renzaglia and Dengate, 1998), Equisetum hyemale (Duckett, 1973), Onoclea sensibilis (Kotenko, 1990), Asplennium trichomanes (Gori et al, 1997), Lygodium japonicum (Liu et al, 1999), Phyllitis scolopendrium (Muccifora et al, 2000), Zamia intergrifolia and Ginkgo biloba (Mizukami and Gall, 1966; Gifford and Lasor, 1980) have been studied. In pteridophytes, the development of blepharoplast in Pteridium aquilinum was investigated in detail by Bell (1974a) and Bell and Ducket (1976). Hepler (1976) studied the origin and development of the blepharoplast of Marsilea vestita in detail. Another organelle that arises de novo after the blepharoplast is the multilayered structure. Carothers and Kreitner (1967) reported a new organelle called Vierergruppe in spermatid of Marchantia polymorpha, its outer layer, called spline, is composed of a few tubules, which lie parallel and close to each other. The second and third layers under the spline consist of strata, and the undermost layer is also composed of parallel tubules. Norstog (1974) and Gifford and Lasor (1980) described a similar structure to the Vierergruppe in Z. intergrifolia and G. biloba. Duckett and Bell (1969) replaced the Vierergruppe by the MLS for the first time in the fern Eqisetum. The MLS of Eqisetum, which resemble the Vierergruppe of the M. polymorpha, also consists of four layers. Myles and Hepler (1977) considered that the MLS of Marsilea vestita is composed of three layers: the outer layer of the MLS is microtubular ribbon (MTr); the second layer is partition, and the inner layer is a dense plaque. Kotenko (1990) states that the MLS of O. sensibilis only consists of MTr and its inner lamellar plates. The MLS of A. trichomanes (Gori et al, 1997) and P. scolopendrium (Muccifora et al, 2000) is similar to that of O. sensibilis. It is noteworthy that the blepharoplast and the MLS form and develop in spermatogenesis of the archegoniates. The blepharoplast and MLS belonging to different groups of plants are quite different. It is now widely recognized that clarification on phylogenetic relationships between different groups of plants could be achieved through consistent studies of blepharoplast structure and development. Up to now, papers on blepharoplast and the MLS of the protoleptosporangiopsida have not yet been published. We choose the primary fern Osmunda cinnamomea var. asiatica as material to research the ultrastructural features of the blepharoplast and the MLS. 1 MATERIALS AND METHODS The spores of Osmunda cinnamomea var. asiatica Fernald identified by Prof. BAO Wen Mei of Harbin Normal University were collected in late May 1999 in Mount Laoshan, Shangzhi County, Heilongjiang Province, China. The fresh spores were sown in the modified Knop’s culture medium by using the method of Bao (1985). At room temperature and continuous illumination with fluorescent light, the densely sown spores in the culture dishes could form male gametophytes in about five weeks. At this time, some antheridia developed on the margin or on the lower surface of the gametophytes. The gametophytes in different developmental stages were selected and plunged into 5% glutaraldehyde in 0.1 mol/L phosphate buffer at room temperature for 7 h. The material was subsequently washed with the same buffer and fixed in osmic acid (2% aqueous) for 2 h at room temperature. The fixed material was also washed with the buffer and dehydrated in a graded acetone series, then embedded in Epon 812. Specimens were thin-sectioned with a new-made glass knife on an LKB-Nova ultramicrotome. The thin-sections were stained at room temperature with uranyl acetate and then lead citrate. All specimens were observed and photographs taken under a JEOL-2000 electron microscope. 2 RESULTS The spores of O. cinnamomea var. asiatica sown densely in the culture medium form narrow heart-shaped gametophytes, i.e. prothallus. Some cells in the margins and/or lower surface of the prothallus differentiated into the initial cells of the antheridia. These initial cells, whose volumes are smaller and cytoplasm is denser than those of the vegetative cells, form rounded antheridia through several mitoses. The antheridia connect with the prothallus through the basal cell. The jacket cells of the mature antheridia consist of a cap cell and seven annular cells (Fig. 1). The inside of the sterile jacket cells is a spermatogenous tissue which is derived from a spermatogenous cell. The spermatogenous cell forms 16 spermatocytes through four mitoses. The 16 spermatocytes produce 32 young spermatids at the last division (Fig.2). Spiral spermatozoids are formed by the differentiation of the young spermatids. The cells in the antheridium are polygonal before the spermatocyte formation. The blepharoplast is not detectable in the polygonal cells until the young spermatids, formed by the last division of the spermatocyte, are beginning to become spherical (Fig.2). The blepharoplasts, belonging to different developmental stages, can be seen near the nuclei of these spherical cells (Figs.3, 4). The differentiating blepharoplast, with a diameter of about 1.0 µm, is a spherical body, and its center is composed of densely stained granular material, i.e. osmiophilic material and some cylinders appearing outside the center. The microtubules can be seen within the osmiophilic material (Fig. 3). The differentiated blepharoplast, also spherical with about 1.2 µm diameter and no densely stained material in the center, consists of scattering or radially arranged cylinders (Figs.5, 6). The cylinders, about 150 nm in diameter and 300 nm in length, are subunits of the blepharoplast (Figs.5, 6). And the cylinder with nine triplets in the outer part has an electron-opaque core, i.e. the axis of the cylinder in the center, from which nine spokes extend to the triplets respectively and exhibit cartwheel-pattern in cross section (Fig.5). Sectioned longitudinally the cylinder displays a columnar profile, in which the hub can clearly be seen (Fig.6). As shown through the successive sections, it is estimated that the blepharoplast contains 30 cylinders. One or two well developed mitochondria and some endoplasmic reticulum can be seen in the cytoplasm near the blepharoplast (Figs.4, 5), as well as some radially arranged microtubules distributed around it (Fig.6). And there are also some microtubules along the surface of the nucleus (Fig.4), all of them seem to extend into the blepharoplast. Along with the further development of the spermatid, the channels between young spermatids disappear and the spermatid begins to differentiate independently (Figs. 7, 12, 13). At this stage, the MLS appears in the spermatid when the blepharoplast is completely differentiated, and a mitochondrion becomes associated closely with the inside the especially at their junction with the microtubular ribbon, which is the outer layer of the MLS (Figs.7, 8). The short columnar cylinder, whose longitudinal axis is at an angle of about 30-40°C to the MTr (Figs.7, 13), extends towards both ends, generating the axoneme of the flagella in the distal end and the wedge-shaped structure in the proximal end, respectively (Figs.8, 9). The axoneme has the typical “9+2” pattern in cross-section and extends out from the plasmalemma (Figs.8, 9). The densely stained plate between the basal body and the axoneme is a terminal plate which is associated with the plasmalemma (Fig.8). The wedge-shaped structure, with a length of about 1.2 µm, is almost four times larger than the basal body, and connect with the MTr. The inner structure of the basal body arranges newly along with the development of the basal body. At this time, the basal body can be divided into two parts. The upper part with a length of about 450 nm from the terminal plate to the middle part has no hub in the center (Fig.8) and exhibits a “stallae pattern” (Fig.9). The lower part with a hub in the center and a length of about 400 nm exhibits a “cartwheel pattern”(Fig.9). The end of the wedge-shaped structure connects with the MTr (Fig.8). In the developmental stage of the blepharoplast, there are channels between the young spermatids, each of which contains a large nucleus, and the chromatin condenses gradually (Fig.2). The cytoplasm is provided with active cytoplasmic organelles, especially the Golgi bodies which have well developed cisternae and many vesicles around it (Figs.10, 11). This may indicate that the spermatids are all at the differential stage. The mitochondrion possesses distinct ridges which are developed better than the previous stage (Figs.4, 6). The form of the thylakoid and its strata in the plastid have degenerated. Its structure is indistinguishable. The MLS grows spirally and gradually approaches the nucleus (Fig.13). The rudimental MLS only consists of about 50 lamellar layers (LL) , each of which is parallel to one another and forms a strip whose height is about 90 nm and width about 500 nm (Fig.14). The giant mitochondrion lies on the inside of the lamellar layers (Fig.12). At the same time, a nuclear crest forms on the surface of the nucleus opposite to the MLS (Figs.12, 14). While the rudimental MLS approaches the nuclear crest, a few microtubules, forming in the outside of the lamella layers, seem to extend to the nucleus (Figs.12, 14). The middle developmental MLS is composed of three layers. The outer layer of the MLS is the MTr, which is composed of a single row of microtubules, each of which runs parallel to one another. The middle layer of the MLS is the lamellar layers, which increases in number up to 70 and with the same height as that of previous stage and width of about 700 nm broader than that of previous stage. Under the lamellar layers, a densely stained layer formed newly which referred to as plaque (Fig.15). The nuclear crest, then, becomes more prominent just opposite to the lamellar layers (Figs.15, 17). The completely differentiated MLS appears in the mature spermatid. It makes approximately 1.5 gyres in the anterior region of the mature spermatid (Fig.23). At this time the MLS forms an accessory band and osmiophilic crest out of the MTr and a layer of osmiophilic material, which surrounds the giant mitochondrion (Fig.16). There is densely stained clump of osmiophilic material which lies in the upper of the osmiophilic material layer (Fig.16). At this stage the MLS and the condensed nucleus are connected by the MTr and the distance between them is elongated (Figs.16, 23). The MLS after its formation is almost changeless in cross-section, but can elongate continuously in longitudinal direction and grows spirally. When the MLS approaches the nucleus, the MTr grows out from the MLS rapidly and extends onto the surface of the nucleus (Figs.15, 17). The MTr increases its number in cross-section (Figs.17, 18) and grows continually along the nuclear surface in a longitudinal direction (Fig.20). The oblique sections indicate that the long axis of the MTr lies at an angle of about 40o to the long axis of the lamellar layers (Fig.19). Along with the extending of the microtubules on the surface of nucleus, the nuclear envelope under the MTr specializes. The two membranes of the nuclear envelope become distinct, lie parallel to each other and are devoid of nuclear pores (Figs. 20, 21). But it is difficult to distinguish the two membranes of the nuclear envelope where they are devoid of the MTr (Figs.13, 22). The MTr is very closely pressed against the outside of the nuclear envelope and no gaps are seen both in cross and longitudinal section (Figs.20, 21). The structure has been identified as the microtubular ribbon-nuclear envelope complex. Finally, the MTr becomes a microtubular skeleton running over the spermatozoid. In the developmental stage of the MLS, the distribution of the basal bodies increases on the outside of the MTr (Figs.17, 20). At the same time, the chromatin is condensed further and the volume of the nucleus becomes smaller than at the previous stage. The active Golgi body and numerous vesicles that contain the half electron-opaque or granular material can be seen in the cytoplasm (Fig.22). These vesicles are continually released from the plasmalemma. In addition, the MTr is also closely associated with the plasmalemma. Therefore, the plasmalemma is considered as a part of the complex. The formation of the complex squeezed the cytoplasm between the nuclear envelope and the plasmalemma into the posterior region (Fig. 17). Finally, a spiral spermatozoid with a condensed chromatin nucleus is formed in the spermatid (Fig.23). 3 DISCUSSION 3.1 The occurrence and function of the blepharoplast and its subunitsFerns, bryophytes and part of the gymnosperm (such as Cycad and Ginkgo) all produce a new organelle, i.e. the blepharoplast. The blepharoplast, a densely stained spherical organelle, generally appears in the spermatocyte and is interpenetrated by numerous lightly stained cylindrical channels (Helpler, 1976; Vaudois and Tourte, 1979; Kotenko, 1990). As to the origin of the blepharaplast, Vaudois and Tourte (1979) studied the motile apparatus of Pteridium aquilinum and considered that the blepharaplast was derived from the dense electron-stained granular material. The blepharoplast of Marsilea vestita has been studied by Hepler (1976). It arises twice in the dividing spermatogenous cells. First it is a flocculent material which soon degenerates; and then arises again close to an indentation of the nuclear envelope and the newly formed blepharoplast also consists of flocculent material with two plaques. Taking the two plaques as the pole, the blepharoplast divides into two. Therefore, it was postulated that the nucleus took part in the formation of the blepharoplast (Helpler, 1976). The differentiating blepharoplast is seen in the young spermatid of O. cinnamomea var. asiatica. The undifferentiated granular material in the center of the blepharoplast indicated that the blepharoplast is derived from the densely stained granular material or the flocculent material. Our observation is similar to that of Vaudois and Tourte (1979) and Hepler (1976). But the indentation in the nuclear envelope was not seen in the spermatid of the O. cinnamomea var. asiatica. The nature and function of the blepharoplast are one of the issues in which most authors are interested. As the blepharoplast differentiates the subunit such as the centrioles or basal bodies and even the MLS, some authors have concluded that it is the microtubule-organizing center (MTOC) (Helpler, 1976; Myles and Hepler, 1977; Liu et al, 1999). In recent years, some authors studied the blepharoplast using the immunofluorescence technique and anti-tubulin antibody identification. Thus, Doonan et al (1986) reported that the blepharoplast of the fern Platyzoma microphyllum contains tubulin at all stages of development. Hoffman and Vaughn (1995) used the Centria as the antibody to label the spermatid of the fern Ceratopteris richardii, and found that the blepharoplast was associated with a quantity of antibodies. These results indicated that the blepharoplast is really the MTOC. The differentiating blepharoplast of O. cinnamomea var. asiatica forms some cylinders in the outer of it. In addition some microtubules can be seen within the granular material and the cytoplasm around the blepharoplast. The observations here in general are similar to those of previous authors and support the view that the blepharoplast is the MTOC. The blepharoplast forms its subunit, i.e. the cylinder, rapidly. The cylinder of O. cinnamomea var. asiatica is composed of a hub, nine spokes and nine triplets that exhibit the cartwheel structure in cross-section. It is equivalent to the centrioles of E. hyemale (Duckett, 1973), the basal body of M. vestita (Myles and Hepler, 1977) and the prebasal body of G. biloba (Gifford and Lasor, 1980). Whatever the subunits of the blepharoplast are called, they actually are the precursor of the basal body and finally become that structure which then produce the flagellum. The cylinder of O. cinnamomea var. asiatica with a diameter of about 150 nm and a length of about 300 nm, is just identical with the centriole of the E. hyemale (Duckett, 1973) and the cylinder of the P. aqulinum (Bell and Ducket, 1976) in size, but much shorter than the basal body of Zamia, whose length almost reaches 4.0 µm (Mizukami and Gall, 1966). As the cylinder change into the basal body, its inner structure and length are also changed with it. The length of the elongated cylinder of O. cinnamomea var. asiatica almost reaches 1.2 µm, equivalent to that of E. hyemale (Duckett, 1973) andM. vestita (Myles and Hepler, 1977), but also much shorter than that of Z. integrifolia whose elongated basal body is 35 µm long (Norstog, 1967). It seems that the length of the cylinder and the basal body may be related to its evolutionary status and may reflect the differences in phylogenesis between different groups of plants. The number of the blepharoplast subunits in the fern is the same as that of the flagella of the same fern (Mizukami and Gall, 1966; Duckett, 1973; Helpler, 1976). Based on the observations under the light microscope, Bao et al (1985) found out that the flagellum number of the spermatozoid of O. cinnamomea var. asiatica is about 28, which is close to the number in the present observations (about 30 subunits) by successive sectioning. Yuasa (1935) counted the sperm flagella of some pteridophytes, in which we found that the flagellum number of the primary species is under 40. But the flagella of Zamia reaches almost 20 000 (Norstog, 1967). The flagellum number of O. cinnamomea var. asiatica perhaps also reflects a basic feature of primary ferns. 3.2 The occurrence and function of the MLS The MLS is another important organelle in spermatogenesis of ferns. Most authors have reported that the MLS is derived from the blepharoplast. Bell (1974b) pointed that the MLS of P. aquilinum arises as a localized development of the blepharoplast. Vaudois and Tourte (1979) reported that the blepharoplast of P. aquilinum born two different systems of microtubules: first the centrioles and next the MLS. Duckett (1973) also stated that the MLS is derived from the breaking up blepharoplast. The MLS has not been seen in the blepharoplast of O. cinnamomea, so whether it is derived from the blepharoplast or originates by itself is not yet determined. The typical MLS of O. cinnamomea var. asiatica is composed of three layers which is equivalent to that of the M. vestita (Myles and Hepler, 1977) andP. aquilinum (Bell et al, 1976). But the MLS of E. hyemale consists of four layers. Apart from the MTr and plaque, the lamellar layers can be divided into two parts (Duckett and Bell, 1969; Duckett, 1973). The subsequent authors such as Kotenko (1990), Gori et al (1997), Muccifora et al (2000) reported that the MLS only consisted of two parts in O. sensibilis, A. trichomanes and P. scolopendrium, i.e. the MTr and the lamellar layers. The MLS of O. cinnamomea var. asiatica was observed in the developmental point of view. In early stage, it only consists of lamellar layers. In mid stage, it is composed of three layers, i.e. the MTr, the lamellar layers and the plaque, which is considered to be the typical MLS mode. In late stage, the MLS forms accessory band, osmiophilic crest and osmiophilic material layer. The number of lamellar layer increases along with the development of MLS. Bell (1974) and Bell and Duckett (1976) stated that it should be related to the nuclear crest, because they found vesicles between the MLS and the nuclear crest, but these vesicles have not been seen in the spermatid of O. cinnamomea var. asiatica. The function of the MLS has not been completely recognized. However, it is certain that the MLS is the initial point of spiral differentiation of the spermatid. Through detailed studies on spermatogenesis in various archegoniates, even in algae (Myles and Hepler, 1977; Duckett et al, 1979; Graham and Mcbride, 1979; Miller and Duckett, 1983), there is a agreement that the MLS or the lamellar layers are an elaborate MTOC, which is responsible for the ordered assembly of the microtubules. It is proposed that the microtubules, as important components of the cytoskeleton, exist in all kinds of cells. But, their distribution and arrangement exhibit a certain flexibility. Only with the MTOC, can the microtubules be arranged in order. In the present study, the lamellar layers of O. cinnamomea var. asiatica appears before the MTr forms and some microtubules are seen outside of the lamellar layers. It is reasonable to propose that the lamellar layer is the MTOC. In the present study, it was shown that the MTr forms in the development of the spermatid of O. cinnamomea var. asiatica. When the MLS approaches the nuclear crest, the MTr extends onto the surface of the nucleus and grows along the surface of the nucleus. At the same time, the MTr and the nuclear envelope unite structurally as a complex. Bell (1978) reported this structure for the first time in the spermatogenesis of P. aquilineum. Miller and Duckett (1983) used sonication and Triton X-100 to disrupt the mature gametes of two mosses which reveals that the connection between the MTr and the nuclear envelope is extremely firm. The complex is prominent in spermatogenesis of O. cinnamomea var. asiatica. The nuclear envelope under the MTr develops a special form; the two nuclear membranes are parallel to each other and devoid of nuclear pores. Therefore, it is suitable to name it as a complex. However, the MTr is also closely associated with the plasmalemma. It is suggested that the complex should include the plasmalemma. As the spermatid matures, the MTr becomes a unique organelle running through the spermatozoid and connecting the condensed nucleus, the giant mitochondrion and the flagellar band. Finally, a spiral spermatozoid is supplied with energy by the giant mitochondrion and can then swim into the archegonium to accomplish the fertilization process. REFERENCES:
(Managing editor: WANG Wei) Copyright 2003 - Acta Botanica Sinica. Free, full-text also available from http://www.chineseplantscience.com The following images related to this document are available:Photo images[as03019f7-11.jpg] [as03019f18-23.jpg] [as03019f1-6.jpg] [as03019f12-17.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}