 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Revista Científica UDO Agrícola, Vol. 2, No. 1, 2002, pp. 14-21 Cariología de Dos Especies del Género Tabebuia Gomes (Bignoniaceae) Karyology of Two Species of Genus Tabebuia Gomes (Bignoniaceae) Alcorcés de Guerra, Nilda Universidad de Oriente. Núcleo de Monagas. Escuela
de Ingeniería Agronómica. Code Number: cg02003 RESUMEN De acuerdo con Gentry (1982), Tabebuia chrysantha (Jacq.) Nichols., es una especie extremadamente polimorfa y probablemente T. capitata (Bur. & Schum.) Sandw. y T. subtilis sean variantes geográficas y ecológicas de la misma. También podría formar híbridos con T. ochracea; por tales razones este grupo de especies forma un complejo que dificulta su identificación taxonómica cuando se utilizan sólo las herramientas convencionales de la taxonomia clásica; planteándose la necesidad de recurrir a estudios biosistemáticos. Son éstos los antecedentes que condujeron a realizar el presente trabajo, con el objeto de realizar un análisis cromosómico de Tabebuia chrysantha y T .capitata, especies frecuentes en el estado Monagas, Venezuela; la primera prolifera en zonas montañosas, la otra en la región de los llanos, pero muy distantes una de la otra. Se utilizó meristemo radicular para la observación de los cromosomas y se utilizó colchicina al 0,05%, NaCl al 0,03% y Orceína FLP al 2%. Se determinó el promedio de las medidas de los cromosomas, midiéndose a cinco células por especie. Los resultados determinaron que T. chrysantha presenta los cromosomas de mayor tamaño que T. capitata. Las fórmulas cariotípicas propuestas son: T. chrysantha: 2n = 40 = 2M +26m + 12sm y T. capitata: 2n = 40 = 40m; estos resultados parecen indicar que se tratan de especies que deberían estar en grupos diferentes. Palabras claves: Tabebuia, cromosomas, cariotipos, Bignoniaceae. ABSTRACT In accordance with Gentry (1982), Tabebuia chrysantha (Jacq.) Nichols., is a species extremely polymorphic and probably T. capitata (Bur. & Schum.) Sandw. and T. subtilis could geographical and ecological variances of the same one. It could also form hybrid with T. ochracea; for such reasons, this group of species form a complex that hinders their identification taxonomically when the conventional tools are only used by classic taxonomy; putting forward the necessity to appeal to biosystematic studies. These plows the antecedents that drove to carry out the present work, in order to carrying out to chromosomal analysis of Tabebuia chrysantha and T.capitata, frequent species in the state Monagas, Venezuela; the first one proliferates in mountainous areas, the other ones in the region of the plains but very distant one of the other. Radicle meristem, colchicine at 0.05 %, NaCl at 0.03 % and Orcein FLP at 2 % were used for the separation of the chromosomes, measuring five cells per species. The results determined that T. chrysantha presents the chromosomes of greater size than T. capitata. The cariotipic formulas proposed are: T. chrysantha: 2n = 40 = 2M +26m + 12sm and T. capitata: 2n = 40 = 20m; these results seem to indicate that they are different species. Key words: Tabebuia, chromosome, karyotype, Bignoniaceae. INTRODUCCIÓN El género Tabebuia está ubicado dentro de la familia Bignoniaceae en cualquiera de los sistemas de clasificación filogenéticos conocidos. Este género está constituido por aproximadamente 100 especies; para Venezuela (Gentry, 1982) indica 25 especies; mientras que para el estado Monagas Tineo (1994) publica 10 especies. Son especies predominantemente leñosas, de hojas opuestas, simples o compuestas; por otro lado, del análisis de cinco especies que específicamente presentan problemas taxonómicos, se ha encontrado que sus flores son de color amarillo, blanco y de rosado a violeta y rojo; están ampliamente distribuidas desde el norte de México hasta el norte de Argentina. Gentry (1982) contempla a estas dos especies dentro del grupo de T. capitata, por considerar que comparten muchas características. Son pocos los estudios citogenéticos realizados en este género, posiblemente se deba al pequeño tamaño de sus cromosomas, o al ennegrecimiento que sufren las raíces cuando son fijadas. De acuerdo a la bibliografía el número básico de cromosomas para la familia Bignoniaceae es x = 15, 16, 18, 20, 21, 22 y 40; y en particular el género Tabebuia presentael número básico x = 20. Los estudios cromosómicos de las especies ayudan en parte a resolver problemas taxonómicos que ellas pudiesen tener. Por ello se realizó este trabajo, que marca el inicio de una serie de investigaciones con las especies de este género que morfológicamente son muy parecidas y que dentro de una misma especie, se encuentran subespecies que desde el punto de vista de su análisis cariotípico son bien distinguibles. MATERIALES Y MÉTODOS El material biológico
utilizado en este trabajo estuvo conformado por dos especies del género Tabebuia: T. capitata y T. chrysantha, localizadas en el estado Monagas; situado
en la región nororiental de Venezuela a 63°10’44” longitud de Greenwich y
9°44’56” de latitud norte. El material
ubicado de acuerdo con Tineo (1994), fue marcado con pintura, colectado
y prensado, utilizando las técnicas convencionales e identificado con la clave
de Gentry (1982), procediéndose al montaje en cartulinas (para material
herborizado) y depositado en el herbario UOJ ( Herbario Universidad de Oriente
Jusepín), Núcleo de Monagas. La colección del material se realizó en los municipios Maturín y Caripe para las
especies T. capitata y T. chrysantha respectivamente; además del
material para herborizar también se colectaron los frutos en dehiscencia de las
especies citadas, para seleccionar las semillas, las cuales fueron sumergidas
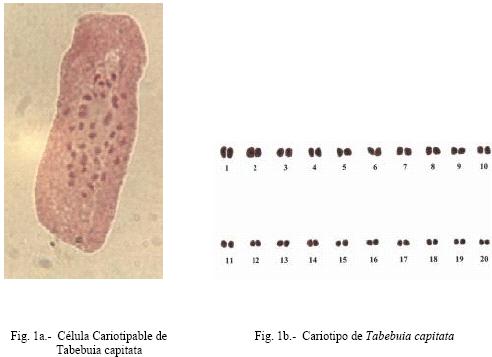
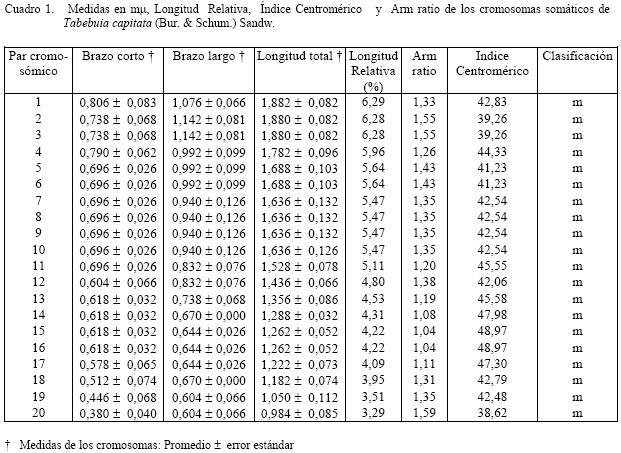
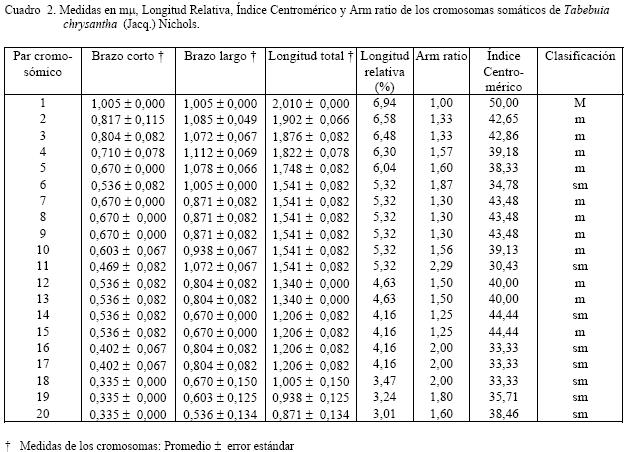
en agua durante A cada una de las especies se les determinó la hora mitótica, calculando en cada caso el número de metafases por campo, para ello se utilizó meristemos radiculares, fijando sus raíces en solución 3:1 (alcohol etílico-ácido acético glacial) durante 24 horas, luego fueron tratadas con ácido clorhídrico al 18% durante 10 minutos para su hidrolización. La coloración fue efectuada con orceína acética al 2%. Una vez determinada la hora mitótica, a 20 meristemos y a diez células por lámina, se procedió al bloqueo de la metafase con colchicina al 0,05%, por un tiempo de dos a tres horas, para luego ayudar a la separación de los cromosomas con cloruro de sodio al 0,03% y colorante orceína acética y orceína FLP (ácido fórmico, ácido láctico, ácido propiónico) al 2% para determinar el número cromosómico de las especies mencionadas. Se analizaron las cinco mejores células por especie que mostraron mas claramente la morfología de los cromosomas, luego fueron medidos con un ocular micrométrico y realizar la conversión a micras, se les calculó la media o medida promedio a cada uno de los brazos y la longitud total de los pares cromosómicos, la longitud relativa, “arm ratio” y el índice centromérico. Con el índice centromérico y el “arm ratio” se realizó la clasificación de los cromosomas de acuerdo a lo establecido por Levan et al. (1964). también se determinó la asimetría intra e intercromosómica, la cual se realizó por los parámetros numéricos, A1 y A2, de acuerdo al método propuesto por Romero-Zarco (1986). Para determinar las diferencias estadísticas a los datos obtenidos en cada una de las especies estudiadas se aplicó la prueba de t de Student con un nivel de significación de α = 0,05. Para la elaboración de los cariotipos se escogieron las mejores células que permitían observar claramente el número y la forma de cada uno de los cromosomas en las especies estudiadas, para ello se fotografiaron y ordenaron de acuerdo a su fórmula cariotípica por parejas de homólogos. RESULTADOS El análisis de los diferentes datos provenientes de las observaciones realizadas determinó que Tabebuia capitata y T. chrysantha (nuevo reporte); presentan un número somático de 2n = 40. Para la especie T. capitata el proceso de germinación de las semillas se realizó en cinco días, con un porcentaje de germinación del 70%. La hora mitótica está comprendida entre las 8:00 – 8:30 a.m. Aproximadamente se prepararon 30 láminas de las cuales se tomaron las cinco mejores en las que se podía apreciar el número y morfología de los cromosomas. El cariotipo propuesto para la especie, consta de 20 pares de cromosomas todos de región media (m) (Figura1.b). Una vez determinadas las medidas de los
brazos cromosómicos se puede decir que los brazos cortos oscilaron entre 0,380 ± 0,040 y 0,806
± 0,083 μm y los largos
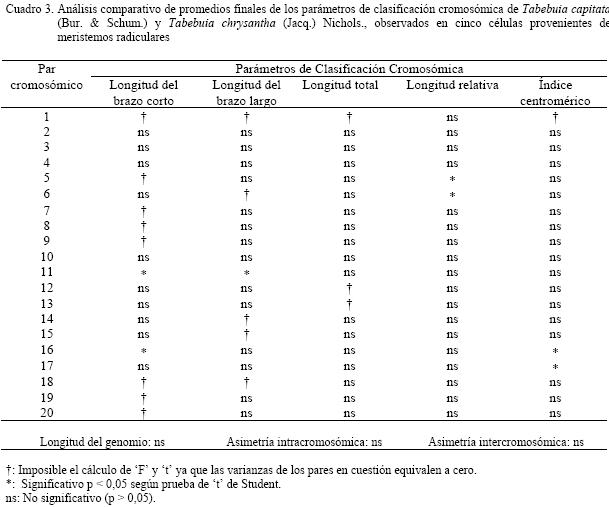
de 0,604 ± Para la especie Tabebuia chrysantha, el proceso de germinación de las semillas fue de siete días y el porcentaje de germinación de un 65%; la hora mitótica está comprendida entre las 5:30 a 6:00 a.m. Aproximadamente se prepararon 35 láminas de las cuales se tomaron las cinco mejores, en las que se podía apreciar el número y morfología de los cromosomas (Figura 2.a). El cariotipo propuesto para la especie, consta de 20 pares de cromosomas, uno metacéntrico (M), el centrómero se encuentra en el punto medio, correspondiente al par uno, 13 pares metacéntricos (m), con el centrómero en la zona media, que le corresponden los pares 2, 3, 4, 5, 7, 8, 9, 10, 12, 13, 14, 15 y 16; y seis submetacéntricos (sm) a los pares 6, 11, 16, 17, 18 y 19. (Fig. 2.b). Una vez determinadas las medidas
de los brazos cromosómicos se puede decir que los brazos cortos oscilan entre
0,335 ± 0,000 y 1,005 ≤ 0,000 mm, los largos de 0,536 ≤ Al realizar el análisis estadístico, se evaluaron longitud del brazo corto, longitud del brazo largo, longitud total de los cromosomas, longitud relativa, el índice centromérico de los veinte pares que conforman el genomio, asimetrías intra e intercromosómica y longitud del genomio entre las especies estudiadas. La comparación entre las especies T.capitata y T. chrysantha, (Cuadro 3), mostró diferencias significativas entre los promedios en: la longitud del brazo corto del cromosoma 11 y del 16; la longitud del brazo largo del cromosoma 11; la longitud total no se apreciaron diferencias significativas; la longitud relativa de los pares cromosómicos 5 y 6; el índice centromérico en los pares 16 y 17. DISCUSIÓN El proceso de germinación en las dos especies estudiadas fue variable, alcanzando un rango de dos a seis días. El porcentaje de germinación fue entre el 75 al 80%. En cuanto a la hora mitótica se apreciaron diferencias significativas entre las especies, con rangos de 5:30 a 10:30 a.m. Es importante señalar que estas especies tienen horas mitóticas muy diferentes, a pesar de compartir caracteres morfológicos fenotípicamente. No hay
información acerca de la hora mitótica de estas especies, sin embargo, Alcorcés
(1994) en el análisis cariotípico de algunas especies del género Capsicum, encontró
que C. annuum var. glabriusculum D’Arcy & Eshbaugh, muestra
como hora mitótica un rango de 5:00 a.m. a 6:00 a.m.; mientras que para C.
annuum var. cerasiforme (Miller) Irish, la hora mitótica oscila de
9:00 a 10:00 a.m. La selección 9 de Capsicum chinense Jacq. muestra como
hora mitótica un rango de 8:00 a. m. a 9:00 a.m. y la selección 10 de la misma
especie la muestra de 6:00 a.m. a 7:00 a.m., Piña (2.001) reporta para la línea
de frijol (Vigna unguiculata) ‘T.C.-9- Como puede observarse la hora mitótica es un parámetro muy variable entre especies y dentro de éstas, es característico para cada especie y más aún para genotipos infraespecíficos, aunque esto no implica que especies diferentes no puedan mostrar la misma hora mitótica. El número cromosómico para las especies estudiadas fue constante (n = 20; 2n=40), lo que corrobora lo citado por Goldblatt (1979-89) y Darlington (1955); considerándose como nuevos reportes el mencionado para T. chrysantha, Los cariotipos de las especies fueron diferentes entre si. Para la especie T. capitata, los 20 pares de cromosomas que conforman el genomio parecen ser muy similares y se clasifican como metacéntricos (m). T. chrysantha, mostró un par de cromosomas metacéntricos (M), donde las diferencias de las longitudes de los brazos corto y largo fue igual a cero, trece pares metacéntricos (m) y seis pares submetacéntricos (sm), donde la longitud de los brazos cortos de los cromosomas están bien marcadas. Es importante señalar las diferencias existentes entre estos dos cariotipos ya que estas especies, de acuerdo a Gentry (1982) muestran muchas afinidades taxonómicas por lo que él las ubicó dentro del complejo T. capitata. Al analizar los
valores promedio de los tamaños de los cromosomas que conforman el genomio, se puede señalar que las
longitudes de los brazos de los cromosomas de T. capitata son más pequeños que los de T. chrysantha.
Aunque no se encontró bibliografía relacionada con la citogenética de este
género, se pueden comparar los resultados de esta investigación con los de
otros géneros. Alcorcés (1997) en un análisis citogenético para las especies Tithonia
diversifolia y Helianthus annuus, indica que los tamaños promedios
son para la primera especie de 4,56 – 8,00 mmy para la segunda
de 6,00 – 10,37μm, estos
cromosomas son de una longitud bastante grande en comparación con los de las
especies de Tabebuia ; Alcorcés (1994) en un trabajo citogenético de
algunas especies del género Capsicum, da a conocer las medidas de los
cromosomas de dos variedades de C. annuum y dos selecciones de C.
chinense, para la variedad glabriusculum los tamaños de los
cromosomas oscilan desde 1,92 - 3,50 mm, para la variedad cerasiforme
los tamaños de los cromosomas van de Las asimetrías intracromosómicas calculadas para las especies estudiadas, coinciden con lo planteado por Romero-Zarco (1986) que cuando los índices se acercan a cero, los cromosomas son metacéntricos. Las asimetrías intercromosómicas o de los cariotipos, es un parámetro que ayuda a visualizar la variación existente entre los cromosomas del cariotipo de una misma especie. Cuando los tamaños de los cromosomas de una misma especie son más o menos uniformes o poco variables entre sí, la asimetría intercromosómica tiende a mostrar valores muy bajos, de acuerdo a los datos obtenidos, los índices de asimetría intercromosómica tienden a mostrar valores muy pequeños, por lo que los cariotipos de las dos especies tienden a ser simétricos. Al analizar el largo total de la cromatina o la longitud del genomio se observó que T. capitata contiene menor cantidad de cromatina que T. chrysantha. Las fórmulas cariotípicas propuestas son diferentes para las dos especies. El análisis estadístico realizado (Cuadro 3) a las especies T. capitata, T. chrysantha, determinó que las mismas deben tratarse como especies bien diferentesya que los análisis realizados así lo demostraron. Gentry (1982), sostiene que la especie T. chrysantha es morfológica y ecológicamente polimórfica y que T. capitata es una variante ecológica de T. chrysantha y además sostiene que ésta especie forma híbridos con T. ochracea. Es importante señalar que las colecciones de estas especies específicamente de T. chrysantha sólo se encontró en el municipio Caripe, las otra especie se encuentra muy distantes en la ubicación geográfica de los sitios de recolección, por lo que se plantea que esas diferencias observadas, podría ayudar en parte a considerar a esas especies como especies separadas y no como un complejo. CONCLUSIONES A pesar de que estas especies se encuentran dentro del grupo de T. capitata como lo organizó Gentry (1982) es importante señalar que T. capitata y T. chrysantha, se encuentran ubicadas geográficamente muy distantes una de la otra y sus características morfológicas son bien diferenciables, además sus cariotipos son diferentes, por lo que se pueden tratar como especies separadas y no como un complejo o grupo. AGRADECIMIENTO Al Consejo de
Investigación de LITERATURA CITADA
The following images related to this document are available:Photo images[cg02003f2.jpg] [cg02003t2.jpg] [cg02003t3.jpg] [cg02003t1.jpg] [cg02003f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}