 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Biomedical Research, Vol. 8, No. 2, 2005, pp.123-125 Short communication Acute Restraint Stress Induces Hyperalgesia via Non-Adrenergic Mechanisms in Rats +*OYADEYI A. S, +AJAO F. O, #IBIRONKE G.F, +AFOLABI A.O Department of Physiology, +Ladoke Akintola, University Of Technology,
P.M.B. 4000, Ogbomoso, Oyo State and #College of Medicine, University
of Ibadan, Nigeria. Received: November, 2004
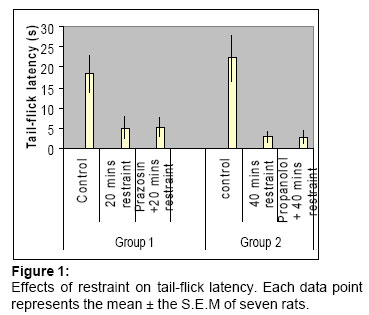
Code Number: md05022 ABSTRACT Analgesia or hyperalgesia has been reported to occur in animals under different stress conditions. this study examined the effect of acute restraint stress on nociception in rats. Acute restraint stress produced a time-dependant decrease in pain threshold; this hyperalgesia was not affected by prior administration of adrenergic blockers suggesting the non-involvement of adrenergic mechanisms. The hyperalgesia may however result from a change in the affective state of the animal and not from a change in sensory processing of noxious stimulus. Keywords:Stress, hyperalgesia, tail-flick test, sympathetic nervous system. INTRODUCTION Humans and animals live in a complex environment and are subjected to various biological and physical stimuli that induce a number of behavioural and physiological responses including alterations in pain sensitivity. An increase in pain threshold following exposure to painful or stressful events was first demonstrated by Akil et al (1976) and Hayes et al (1978). This phenomenon, generally referred to as stress – induced analgesia, has been confirmed repeatedly (Tierney et al, 1991; Marek et al, 1992; D’anci et al, 2000; Koltyn, 2000). As reported by Jorum (1988). However, there is a remarkable paucity of information on stress-induced hyperalgesia due to an in-exhaustive study of the effect of different stressors on nociception. The authenticity of stress-induced analgesia and hyperalgesia is now undisputed but what remains to be fully elucidated is the involvement or otherwise of adrenergic systems in these states. Since the mid 1970s an increasing number of studies have examined the role of adrenergic systems in pain. This interest is due, in part, to clinical studies that emphasized the role of the sympathetic nervous system both in analgesia (Tasker et al, 1992) and hyperalgesia (Levine et al, 1988). By revealing the effect of a stressor on pain, a better understanding of nociception in general may be obtained, and since most pain tests involve some form of restraint on the animal, the first aim of the present study was to examine the effect of acute stress on pain threshold in rats when the stressor was restraint of either short or long duration. The second arm was to investigate the involvement of adrenergic mechanisms in the observed effects. MATERIALS AND METHODSAnimals: All studies used adult male Sprague-Dawley rats (150-200g). They were housed in colony room where light was maintained on a 12-h light 12-h dark cycle with food and water ad libitum. All animals were allowed a minimum of two weeks for acclimatization to the test environment before any experimental manipulations were performed. Drugs: Prazosin (0.15mg/kg) (Pfizer Canada Inc.) and propanolol (0.15mg/kg (Pfizer Canada Inc) were administered i.p in a volume of 0.2ml. Tail-flick test: Nociception was assessed by a modification of D’Amour and Smith’s (1941) tail-flick method , this test was chosen because of the advantages of sensitivity, reproducibility, wide use and experimental convenience. Each animal was gently hand-held while its tail was wholly immersed in water maintained at 50±1ºс. The latency for flicking its tail out of the water (TFL) was recorded with a stopwatch. Restraint stress: Animals were restrained in a Perspex tube measuring 18cm long with 5cm radius, the tube had both ends covered by wire mesh to allow ventilation. This degree of restraint still allowed some grooming of the fore-paws. The first group (n=7) of the animals had their TFL recorded, three days later they were restrained for 20 minutes immediately after which the TFL was taken again. After 5 days they received 0.15mg/kg prazosin i.p immediately before restraint while the TFL was taken again immediately after restraint. The other group (n=7) went through the same procedures except that they were restrained for 40 minutes and received 0.15mg/kg propanolol i.p. Statistical analysis: The paired t-test was used for within-group analysis while the unpaired t-test was used for between group analysis. Statistical significance was accepted at p<0.05. RESULTSEffects of restraint on tail-flick latency: Following 20 minutes of restraint there was significant (P<0.01) reduction of pain threshold in the tail-flick test. Figure 1 shows that the 40 minutes restraint induced a statistically significant (p<0.05) lower pain threshold than the 20 minutes restraint. In both cases the animal’s response was constant; an initial few minutes of violent escape behaviour followed by a period of almost total calm. Effects of adrenergic blockers on tail-flick latency: Prazosin (0.15mg/kg) did not have any statistically significant effect on tail-flick latency when administered immediately before the 20 minutes’ restraint (Figure 1). Propanolol (0.15mg/kg) also did not have any statistically significant effect on tail-flick latency when administered prior to the 40 minutes restraint. DISCUSSION The present study shows that acute exposure to stress in the form of restraint induced hyperalgesia in the tail-flick test. This hyperalgesia was found to be time-dependent being significantly higher with the longer restraint time (40 minutes) than the shorter one (20 minutes). Prazosin, a selective α1 antagonist and propanolol, a β-adrenergic and 5HT1./5HT2 receptor antagonist did not have any effect on the pain threshold implying the absence of adrenergic mechanisms in the observed response. Several investigators have reported analgesia following restraint (Vidal et al, 1982; Vidal and Jacob. 1982) whereas hyperalgesia has been reported once only in the paw pressure test (Pilcher and Brown, 1983), while we report here hyperalgesia using the tail-flick test. Put together, all these suggestion that the nature of the stressor and the test are possible factors involved in the effects of stress on nociception. If however, the nature of the stress is described in terms of duration our results that the nature of the stressor must be of crucial importance in a determining stress-induced changes in nociception fact that the already been stated by Tierney et al (1991). This point is in obvious contradiction to the contention of Jorum (1988) who postulated that it is not the nature of the stressor but the animal’s response during the stress procedure that determines what stress induced changes in nociception would be. According to him, hyper emotionality (agitation and vocalization) by the animal during stress procedure leads to hyperalgesia while calmness by the animal results in analgesia. During the stress procedure all the animals displayed similar responses; an initial period fo hyper emotionality followed by a longer period of calmness. We would expect the calmness to overrule the effects of hyper emotionality, if the view of Jorum (1988) held true, and thus the result should be analgesia. However, all the animals responded with hyperalgesia. Although the involvement of adrenergic mechanisms in nociception has been severally emphasized (Ono et al, 1991: Taskar et al, 1992), our β study did not find any adrenergic mechanisms in the observed hyperalgesia. Even then we know that the absence of evidence is not the same as evidence of absence. This may point to a possible non-sensory effect of restraint stress on nociception. It is possible for restraint to cause a change in the affective state of the animal, represented as fear or anxiety, and may thus enhance the response of the animal to a given stimulus or alter the myriads of CNS modulatory systems to directly alter the afferent input. In conclusion, hyperalgesia to restraint stress via non-adrenergic mechanisms has been shown. Hyperalgesia to stressors such as other vapors. (Hayes et al, 1978) and horizontal oscillation (Hayes et al, 1978). What remains to be full elucidated is whether the effect of restraint stress on nociception is test-dependent. The present study has serious implications for the pain researcher who often has to restrain animals during pain testing procedures. REFERENCES
© 2005 - Ibadan Biomedical Communications Group The following images related to this document are available:Photo images[md05022f1.jpg] |
| |||||||||

{kind=link}