 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
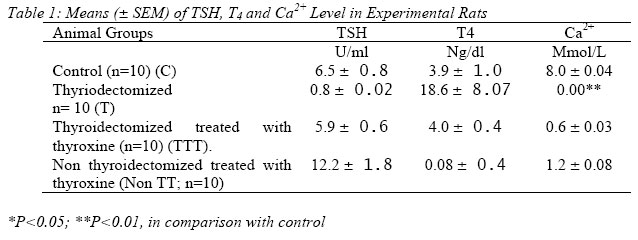
Nigerian Journal of Physiological Sciences, Vol. 20, No. 1-2, 2005, pp. 95-100 DEPENDENCE OF CALCIUM ON THYROID HORMONE FOR THE REGULATION OF CELLULAR FUNCTIONS A. M. KEMALKOLAM,* SABO,* A. B. A ADELAIYE and A. S. SAGAY** *Department of Human Physiology, **Department of Obstetrics and Gynaecology College of Medical Sciences University of Jos, Jos. PMB 2084, Jos Nigeria. Received: 24/9/2005 Code Number: np05018 Summary: Recently, the thyroid hormone has been shown to cause increase in Ca2+ concentration by mobilizing intracellular Ca2+. The mobilization of intracellular Ca2+ in the absence of transmembrane Ca2+ influx has been accepted as evidence for a cell-surface Ca2+- receptor. The possible role of thyroid hormone in the regulation of cellular functions by Ca2+-channel was investigated using parameters proven to indispensably involve Ca2+: Bleeding and clotting times and rat uterine muscle activity in Ca2+-free Tyrode’s solution. Adult female Wistar rats divided into four groups, namely; (i) Control, (ii) Thyroidectomized, (iii) Throidectomized-treated with thyroxine and (iv) Thyroxine-treated; were fed on rat chow. While groups i and ii were allowed access to tap water ad libitum for 60 days, groups iii and iv received tyroxine (10mg/kg body weight) every alternate day and tap water ad libitum for 60 days. All groups were kept in an air-conditioned room till they were euthanized. The results showed that thyroxine elicited biphasic contraction in Ca2+ - depletion and potentiated uterine contraction during Ca2+- loading (P<0.001). Inhibition of contractions in thyroidectomy, showed dependency of Ca2+ on thyroid hormones for the regulation of cellular functions. Key Words: thyroid hormones, Calcium receptor, cellular functions. Introduction Free cytosolic calcium (Ca2+) concentration plays a key role in the regulation of the contraction-relaxation cycle of smooth muscle (Ebeigbe, 1987) and cardiac muscle (Obiefuna, Sofola and Ebeigbe, 1993). Calcium activation of muscle contraction involves Ca2+ binding to the ligand Troponin C and a conformational change of thin filament protein (Ruegg, 1986). Activation is said to be brought about by an increase in Ca2+binding to troponin C and as a result of increased myoplasmic calcium concentration or an enhanced Ca2+ (Marrow and Straton, 1991). The increase in the concentration of extracellular Ca2+ has been reported to activate the Ca receptor linked by G proteins G1 and Gq and by inhibition of adenylate cyclase and stimulation of phosphoilipase C (Siegman, 1987). Transient phasic contractions are elicited by noradrenaline in calcium-free medium, and the calcium pool mobilized by noradrenaline for the contractions has been characterized as membrane bound (Ebeigbe and Aloamaka, 1985; Mecca and Webb, 1984). This agonist-mobilizable membrane bound calcium store in smooth muscle has been demonstrated by electromicroscopy and cytochemistry (Ebeigbe, 1989). The Ca2+ is of limited size and response to the agonist is only sustained by the influx of Ca2+ across the plasma membrane (Karaki et al, 1979). A variety of neurotransmitters and hormones act at surface membrane receptors to activate mechanisms that result in an elevation in the free Ca2+ concentration in the cytosol. This Ca2+ signal is the trigger promoting a range of cellular response (Gallacher, 1988). Calcium enters many processes in haemotology such as haemagglutination, bleeding, clotting and prothrombin times (Baker et al, 1994). Infact there has been reported a calcium receptor dependence for the regulation of cellular functions (Nemeth, 1995). Recently, the thyroid hormone has been shown to cause increase in calcium ion concentration by mobilizing intracellular Ca2+ (Adeniyi et al, 1993; Amadi et al, 1999) for insitu secretion and mobilization of intracellular calcium. The mobilization of intracellular Ca2+ in the absence of transmembrane calcium influx has been accepted as evidence for a cell-surface Ca2+ receptor (Nemeth, 1995). The possible role of thyroid hormone in the regulation of cellular functions by calcium receptors is presently investigated, using parameters proven to involve calcium such as bleeding time, clotting time, prothrombin time and contraction of the rat myometrium; in calcium-free physiological salt solution (De-Jalon’s solution). The rat myometrium, an excitable tissue was chosen to investigate whether such tissue is indeed excitable in the absence of thyroid hormone. It is well known that hypthyroidism leads to hypogonadism. Infertility should be viewed from the couple point of view since management should address the couple as a unit. Investigations in our hospital and research in our laboratory (Amadi et al 1991; 2004; Okeahialam, Amadi and Ameh, 2005) have established that thyroid hormone enhances sperm quality and the contractile responses of the vas deferens. It is therefore imperative to extend this study to the female sex and even the male at the cellular level; moreso on the parameters or factors thought to initiate excitability and therefore cause contraction. Hence the present study. Materials and Methods Animals Sexually mature female albino rats weighing between 250 and 300g were obtained from the University of Jos Animal House, Jos, Nigeria. the rats were divided into four (4) groups of ten (10) rats per group. They were kept in experimental Animal Laboratory to acclimatize for seven (7) days. The rats of all groups were allowed free access to feed and water ad libitum. The ambient temperature of 17oC was maintained by air-conditioning. Thyroidectomy Adult female rats of groups 1-4 (n=5), were exposed to ether aneasthesia. The aneasthetized rats were placed on a thermostacally controlled dissecting table (Bioscience, U.K.), set at 37oC. A modified method described by Adeniyi and Oluwookorun (1989) and Amadi et al, (1996) was followed. The thyroid glands were surgically removed at the upper part of the neck. The wound was carefully swabbed with a disinfectant (70% alcohol) followed by application of procain pecilling solution before suturing. The surgery was carried out under aseptic condition. The operated animals were allowed to recover from the ether aneasthesia and were fed on rat chow and tap water was allowed ad libitum. However, the operated wounds were allowed to heal for 60 days after five (5) days post-operative treatment with operative treatment (5iu/kg). Treatment 40 adult female thyroidectomized Wistar rats were dived into four (4) groups. Group 1, received normal saline as control, group II, thyroidectomized group III, thyroidectomized treated with thyroxine (10m g/Kg; Sigma U.K.), and group IV, non-thyroidectomized treated with thyroxine orally, on alternate days for 60 days. At the end of be treatments, the animals in all the groups were sacrificed and the blood samples were collected through cardiac puncture for preparation of plasma for hormonal assay and uterine muscle strips were carefully isolated and prepared into strips. Hormonal Assay Thyroid Stimulating Hormones (TSH) was measured following double antibody radioimunassay (RIA) kits (Hall et al, 1972). The plasma level of thyroxine of all animals of groups 1-4 were estimated using RIA kits following a modified method of Lee (1964). Uterine Muscle Strip Preparation Thyroidectomised, non-thyroidextomized and treated rats with thyroxine were sacrificed. Their uteri were carefully isolated and placed in a petric dishes containing normal salt physiological solution (Tyrodes solution). uterine muscle strips were prepared from the horns of the uteri of treated and non-treated rats. The preparation was set up in the smooth muscle organ bath system, constantly bubbled with air and the temperature was maintained at 370C. A resting tension of 1.0g was sustained on isometric transducers (Grass FT 03) in 20 ml organ baths each containing Tyrodes solution. The preparation was allowed to equilibrate for 30 min. and the physiological activity of the muscle strips was recorded using Grass Polygraph (Model, 71; Grass Quinay LTD, USA). Determination of calcium flux Calcium flux was ascertained by determining the calcium concentration in the fluid bathing the tissues in each thyroid state. The technique used by Lorenz (1982): Frazer et al, (1986) and Amadi et al, (1999) was applied. Metal complexing dye orthocresolphtalin complexcone (CPC) forms a chromophore with Ca2+ in alkaline solution. The colour produced is measured spectrophtometrically at 575mm. The chromogens reagent contain 8-hydroxyquinoline to mask interfering cations such as Mg2+. Colour intensity developed by the Ca2+ complex increased proportionately with increase in pH and was also dependent on the buffer species employed. Diethenolamine (DEA) was added to enhance the colour intensity and to maintain the pH between 10 and 12. A volume of 1.0ml of organ bath fluid was pipetted out while contraction was being determined. Using spectrophotometer SP6 (Model 450 Pye Unicam, Philips), the calcium concentration was determined by measuring the abosrance (A) of the developed colour. Ethelene diamine tetracetic acid (EDTA) was added to chelate Ca2+, to correct for haemolysis and to dissociate the Ca2+ chromogen complex but the Hb-chromogen complex remained undissociated. The absorbance due to the Hb-chromogen complex was subtracted from the total absorbance to give Ca2+ absorbance. Orthocressolphtalein complexone was used for the assay. Determination of Bleeding Time Bleeding or clotting time was estimated in the four groups of rats; thyroidectomized and non-thyroidectomized treated and non-treated with thyroxine rats, respectively, following a modified method of Duke (1994). A tail vein of each of the rats was punctured. The time of puncture and the first appearance of the blood was noted. A piece of filter paper was blotted with the blood without pressing the wound. This was repeated every 10 sec until no blot appeared on the filter paper. The time was then recorded as the bleeding time. Statistical Analysis Statistical analysis was performed using a Aristo M. 800 programmable calculator (Germany). Mean, Standard Deviation (SD), Standard Error of Mean (SEM) were calculated for all sets of data for five or more values. Two sets of data were compared by student’s t-test. Probability values (P-values) given in the text refer to student’s t-test unless specifically stated that they refer to chi-square of paired t-test. Results Figure 1 shows the magnitude of Thyroxine induced phasic contraction during calcium loading. The dose response effects of calcium chloride were potentiated on the muscle strip activities of the thyroidectomized treated with thyroxine (TTT) and non-thyroidectomized treated with thyroxine (TT) and shifted the curves to the left (fig. 1; P<0.01 and 0.05 respectively). On the ohter hand thyroidectomy (T) inhibited Cacl2 induced dose response effects and shifted the curves to the right side of the graph (fig. 1; P<0.01). The hormonal assay results showed that thyroidectomy caused a significant decrease in the plasma level of thyroid stimulating hormone (TSH), P<0.01, and calcium (Ca2+), (P<0.01). Ironically, significant increase in the plasma concentration of thyroxine was observed, (P<0.01; table 1). Thyroidectomised rats treated with thyroxine (TTT) reversed the reduction plasma concentration of TSH amd Ca2+ by raising TSH and Ca2+ plasma concentration respectively. Synergistic release of T4 following thyroidectomy was inhibited by thyroxine treatment (table 1). Normal rats (non thyroidectomized rats) treated with thyroxine showed increase plasma level of TSH by 100% compared to control, as well as decrease plasma levels of T4 and Ca2+ respectively, (P<0.05; table 1). Effect of Thyroxine Thyroxine (10-4 – IM) induced contractile responses of the uterine muscle strips in a dose related manner (Fig. 1). The contractile responses in the myometria were the indication of the ca2+ mobilization from muscle membrane bound pool. The uterine muscle strips (3cm long) were set up in 20ml baths containing Thyrode’s solution constantly bubbled with air. Bleeding Time Table 2 summarizes the bleeding clotting and prothrombin times for the groups of 10 rats each. Thyroidectomy significantly prolonged the bleeding time, clotting time and prothrombin time (P<0.001). Thyroxine treatment within 60 days did not significantly change the parameters when compared with control and thyroidectomized rats-treated with thyroxine. Discussion Thyroxine treatments has been shown to cause potentiation of contractility by increasing calcium concentration (Adeniyi et al, 1992, 1993; Amadi et al, 1996, 1999; and Prostaglandin E2 itself has been reported to evoke Ca-release in situ for contraction (Burket et al, 1999). Presently, in Calcium-free physiological, salt solution (PSS) thyroid hormone (T4) has been shown to elicit biphasic contractions, which are proportionate to increasing calcium activation of its mobilization. This mobilization of intracellular calcium in the absence of transmembrane Ca2+ influx as observed in the Ca2+-free PSS in this study has been accepted as evidence of a cell-surface Ca2+ receptor (Ebeigbe, 1989; Nemeth, 1995,). The increasing T4 concentrations were shown to also increase Ca2+ - contractions during Ca2+ loading. This observation is further confirmed by the results of the present investigation that thyroidectomy suppressed the intracellular Ca2+ by inhibiting its activation for the contraction of the myometrium, and by prolonging Bleeding Time. It is thus the secretion of Ca2+ regulated by the level of thyroid hormones that modulates cellular activity. Calcium channel activation rather than the associated increase in Ca2+ concentrations is the key factor regulating secretion (Nemeth and Scrapa, 1986). Thyroid hormone has been shown in this study to be an agonist of calcium by showing biphasic contractions and a mobilizer of intracellular Ca2+-free PSS and also a primary activator of Ca2+ receptors for the potentiation of myometerial contraction. The calcium pool mobilized by T4 for phasic contractions in Ca2+ - free medium PSS in this work is membrane bound. Ebeigbe Aloamaka have shown that noradrenaline mobilizes membrane-bound calcium in producing phasic contractions. Thyroid hormone from this work could be suggested to be calcimimetic which could provide antiresorptive therapy by inhibiting calcium loss. T4 thus targets calcium receptors in cells such as smooth muscle of the myometrium and red blood cells, and might provide novel therapies for some diseases. It might find utility in bone and mineral disorders, especially if it is counter-balanced by an appropriate calcilytic drug or agent. Bleeding or clotting time showed prolonged, abnormal values in thyroidectomy. The values in thyroxine treatment and in the thyroidectomized group treated with thyroxine fell within normal values when compared with controls. This was probably as a result of depressed calcium mobilization and thus its inactivation as observed by Adeniyi et al, 1993 and Amadi et al, 1999. Bleeding or clotting time is achieved only in the presence of Ca2+ among other haemostatic factors (Baker et al, 1994). This calcium mobilization by thyroid hormone observed in thyroxine treatment in this study took place in the absence of extracellular Ca2+ influx which is an evidence of cell surface Ca2+ channels reported by Nemeth (1995). Thyroid hormone could be said to target calcium channel for activation in cells such as smooth muscle of the uterus, and red blood cells and could provide antiresorptive therapy by inhibiting calcium loss. It might be suggested to provide novel therapies for some diseases and might find utility in bone and mineral disorders. This study observed that T4 activated Ca2+ - receptors with a result of increased Ca2+ concentration responsible for cellular functions. There appears to be a thyroid hormone dependence of calcium ions concentration for the regulation of cellular functions. Acknowledgement The authors deeply appreciate the services of Mr. Godwin Njoku of Medical Illustration and Training Unit (MITU) for reproducing the figures and Miss Fumilayo God’s Gift Business for Secretariat Expertise. References
© Physiological Society of Nigeria 2005 |

{kind=link}
{kind=link}
{kind=link}