 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
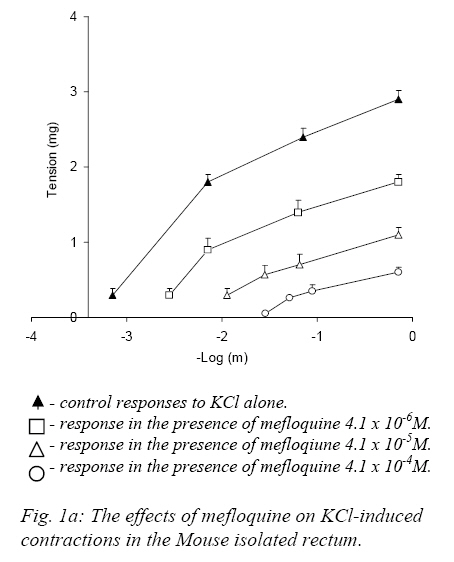
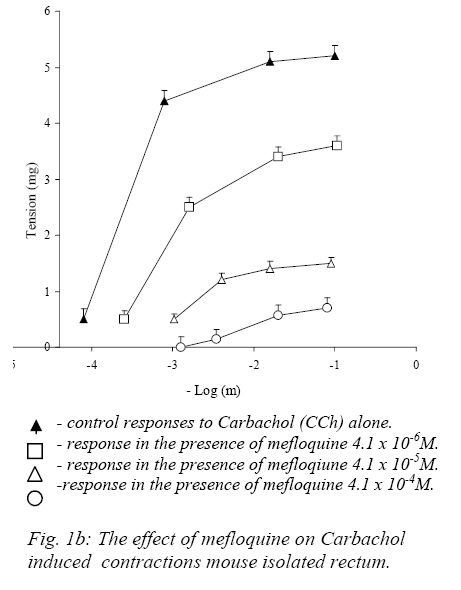
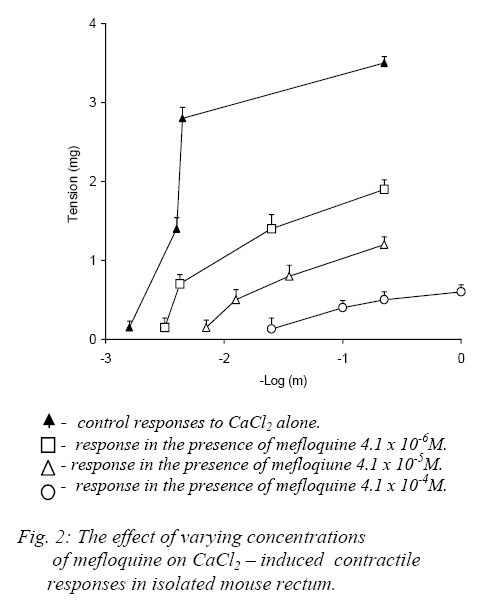
Nigerian Journal of Physiological Sciences, Vol. 22, No. 1-2, 2007, pp. 43-47 Effect of Mefloquine On the Mechanical Activity of the Mouse Isolated Rectal Smooth Muscle P. C. Unekwe, J. O. ogamba, K. C. Chilaka, and J. C. Okonkwo* Department of Pharmacology and Therapeutics, College of Health Sciences, Nnamdi Azikiwe University, Nnewi, Anambra State, Nigeria. Accepted: 2/10/2006 Code Number: np07007 Summary The effects of mefloquine on the mechanical activity of the mouse isolated rectal smooth muscle was studied. Mefloquine (4.1x10-5 - 5.2x10-3M) when applied alone and separately exerted variable effects on the rectum. In some preparations, it caused slight phasic contractions while in others no response was elicited. When the external (Ca2+) was increased from 1.8mM to 300mM mefloquine produced phasic contractile activity which was abolished on return to normal 1.8mM suggesting that the contractile activity was due to extracellular Ca2+ influx. Meflaquine (4.1x10-6M – 4.1x10-4M) caused contraction – dependent inhibition of KCL, Carbachol and CaCl2 (in depolarizing Tyrode Solution). Mefloquine (2.1x10-4M) blocked KCL, but not carbachol contractions which were largely reversed by increasing (Ca2+). The results show that mefloquine possesses anticholinergic and appreciable calcium channel blocking activity. Key Words: Mefloquine, mouse rectal muscle, contraction. Introduction Malaria is a tropical disease which remains an enigma. Despite the best efforts of the medical and pharmacological communities, malaria poses an undiminished threat to people living in or traveling to endemic areas such as Africa. Plasmodium falciparum, the most clinically significant causative organism has demonstrated an unusual propensity to acquire resistance to antimalarial therapy (Bruce-Chwatt 1985, WHO 1986). In Africa alone between 1 and 2 million children die from the disease each year. These figures are almost certainly an under estimate because of the nature of the target population and problems with under-reporting. Indeed, it has been estimated that malaria may occur in about 500 million cases and at least 2.3 million of these will be fatal (Sturchler, 1989). The unprecedented spread of chloroquine-resistant strains of P. falciparum has several weakened the range of drugs available to treat the disease and has increased interest in newer agents such as mefloquine. Mefloquine hydrochloride is DL-erythro-(2-piperidyl)-2-8-bis (trifluoromethy)-4-quinoline methanol hydrochloride). The molecular weight of mefloquine hydrochloride is 414.8 and that of the bases is 378.3. It is an odorless, bitter, white powder, insensitive to beat, light and oxygen even in solutions. It is highly active blood schizonticide against multidrug-resistant falciparum malaria. The effectiveness of mefloquine is essentially a result of destruction of the asexual blood forms of the malarial parasites in humans (Harkanson et al, 1990). Quinine and chloroquine, although mainly known as antimalarial agents exert a number of effects on muscle mechanics. The effects of quinine on skeletal muscle has been extensively investigated (Isaacson and Sandow, 1967, Isaacson et al 1970, Huddert, 1971, Huddert and Price, 1976). Also Ebeigbe and Aloamaka (1982) studied the actions of chloroquine on the mechanical activity of the rat portal vein and showed that chloroquine depressed spontaneous as well as acetylcholine-induced contraction of the rat ileum which they attributed to inhibition of Ca2+ from an acetylchaline-sensitive “membrane-bound” pool. And in 1990, Unekwe et al (1990) studied the effects of chloroquine on the rat urinary bladder strip. The present study was designed to investigate the actions of mefloquine hydrochloride on smooth muscle responses of the mouse rectum with a view to elucidating the mechanism of action of this drug on this tissue. Materials and Methods Healthy young adult Swiss male albino mice weighting between 19-32g were stunned and bled and the whole rectum 1-1.6cm, was dissected from the animal. Each rectum was placed in a petridish containing Tyrode solution and the rectal contents were gently flushed out with Tyrode solution using a 10ml pipette. Each preparation was suspended vertically under a testing tension of about 1g in 10ml organ bath containing Tyrode solution at 37oC and connected to the thread attached to a fixed pin in the tissue bath and the other to a lever writing on a smoked drum. The load on the level was about 1.6g giving a magnification of 8-10 folds. The composition of normal Tyrode solution was (mM/L), NaCL: 138, KCL: 5.7, CaCl2: 1.8, NaHCO3: 2.5, Na2PO4:0.3 MgSO4: 1.1, d-glucose: 5, PH 7.8. The Tyrode solution was continuously bubbled with air and an equilibration period of 60mins usually preceded drug application was employed. During this period, the Tyrode solution was replaced at 10mins intervals to prevent accumulation of metabolites. Contractile responses magnified 8-10 folds were recorded isotonically on smoked drum. In this set of experiments, submaximal concentrations (about EC75-85) of KCL (5.4x10-2M) and CCh (5.8x10-7M) and the concentrations of Mefloquine (4.1x10-4M) which produced 90-100% inhibition of KCL or CCh response were employed. Control KCL or Carbacol response was evoked first in the absence and then in the presence of state concentrations of mefloquine in normal (1.8mM Ca2+) Tyrode solution. Thereafter mefloquine was presented in the experiments. The external Ca2+ concentration (Ca2+) was then increased, and the KCL or carbachol responses determined in the presence of increasing (Ca2+) concentration. CaCl2 responses were determined in Na+-free, Ca2+-free high K+(120mM) depolarizing Tyrode solution. Experimental Protocol Consecutive concentration-response curves were constructed for carbachol (CCh) or potassium chloride (KCL) until the reproducibility of the responses were established and then determined in the presence of varying concentrations of Mefloquine. An agonist drug, carbachol-contact of 60secs was allowed and 5mins drug cycle was employed. Mefloquine was prepared in the Tyrode solution to achieve the desired concentrations and was allowed to equilibrate with the tissues for about 15mins before agonist-induced responses were determined. At the end of such an experiment the tissue was washed for 30-60mins in atropine-free (antagonist) Tyrode solution and agonist-induced responses were again determined in order to assess whether the effects of the atropine sulphate were removable by washout. In some experiments, the effects of mefloquine (agonist) on contractile responses induced by CaCl2 was examined, the tissues were equilibrated for 30mins in a modified Tyrode solution which then contained high potassium concentration (120mM) K+ contraction response curves were then determined in the presence of varying concentrations of mefloquine and again 30-60mins after washout of the antagonists. In these experiments only one antagonist (atropine sulphate) was exposed to each tissue preparation. In another set of experiments, the agonist action of mefloquine was investigated in six preparations, the effects of a range of concentrations of mefloquine was determined. The drug was added cumulatively at suitable intervals and each drug concentration allowed to eliciting its maximal effect. These experiments were performed both in normal Tyrode solution and in modified Tyrode solution containing high (300mM) external (Ca2+). In some preparations of the mouse the reversibility of mefloquine induced-blockade of KCL or CCH contractions by increasing external (Ca2+) was studied. Sub-maximal (60-80%) contractions of KCL or CCH were evoked until reproducible contractions were obtained and then in the presence of a concentration of mefloquine that produced (90-100%) inhibition of KCL or CCH contraction in normal (1.8mM Ca2+) – Tyrode solution. This procedure was then repeated in varying concentrations of external (Ca2+) from 1.8 – 400mM (Ca2+). Mefloquine was prepared in the Tyrode solution at the desired concentration (4.1x10-4 M). at the end of such an experiment, mefloquine was washed out with normal (1.8mM Ca2+)-Tyrode solution for 30-60mins and the sub-maximal KCL or CCH responses were again recorded on the smoked drum (paper) to determine whether the effects of the antagonists, (atropine sulphate, tetrodotoxin and nesostigmine) were removed by washing out. Drugs and Statistics The following drugs were used: Atropine sulphate, Tetrodotoxin, Carbachol chloride and neseostigmine obtained from British Drug House, mefloquine hydrochloride from Hoffmann-La Roche Ltd, Basel, Switzerland. Carbachol was dissolved in 0.1m HCL at high concentrations and dilutions were made in distilled water. All other compounds were dissolved in distilled water. Also all the reagents employed were of analytical grade. Drug concentrations refer to final bath molar concentrations. Results are presented as mean + standard error of the mean (Mean + SEM). Differences between mean values were assessed using the student’s “t”-test for statistical analysis. Values of (P<0.05) were regarded as significant. Results Effects of Mefloquine on Potassium and Carbachol-induced Responses of the Mouse Rectum KCL concentrations (1.3x10-2-1.1x10-1M) and carbachol concentrations (5.5x10-8-5.0x10-6M) each produced concentration – dependent contractions (seen on the traces on smoked paper) of the mouse rectum. The mean EC50 (concentrations that produced 50% of the maximum response) are presented in table 1 Mefloquine (4.1x10-6 – 4.1x10-4M) antagonized in concentration-dependent manner, the contractile responses induced by KCL and carbachol (Fig.1a, b). The blockade produced by the mefloquine in each instance was non-competitive since the agonist concentration response curves were displaced to the right in non-paralled fashions with depressed maxima. The blockade produced by mefloquine was substantially removed by washout between 45-60mins. Carbachol but not KCL or CaCL2 induced contractions were abolished by atropine sulphate (1.4x10-7M). Effects of Extracellular calcium concentration (Ca2+) on mefloquine induced blockade of carbachol and KCL concentrations. Increasing the (Ca2+) produced complete (100%) reversal of mefloquine induced blockade of KCL evoked contractions, whereas, the maximal reversal of mefloquine depressed carbachol contractions recorded was 10.2+4.8%. The difference in the degree of reversal obtained between the mefloquine depressed KCL and carbachol contractions was highly significant (P<0.05). In Na+-free, Ca2+-free high K+ (120mM) depolarizing Tyrode solution, application of CaCL2 (4.4x10-2–4.4x10-1M) caused concentration-dependent reproducible contractions. Mefloquine [4.1x10-6M– 4.1x10-4M] concentration-dependently blocked CaCL2 -induced contractile responses (Fig.2). The blockade produced by mefloquine could be removed by washout after (30-60mins). Substantially the same results were obtained when atropine sulphate, 1.4x10-6M was present in the modified depolarizing Tyrode solution employed in this set of experiments. Table I: EC50 +SEM values of carbachol, potassium and calcium chloride on the mouse isolated rectum.
Discussion The results presented in this study show that mefloquine inhibit the contractility of mouse rectal smooth muscle. Mefloquine produced marked inhibition of the contractile responses evoked by various pharmacological interventions such as carbachol, potassium chloride and calcium (in depolarizing Tyrode solution)-respectively. This suggests that this compound may not be specific in its inhibitory actions on mouse rectal smooth muscle. The mouse rectum preparation exhibits intrinsic (spontaneous) contractile activity of varying degrees. This activity has been previously shown to be entirely neurogenic in origin, since they were abolished by low concentrations of tetrodotoxin (Unekwe and Savage, 1991). Furthermore, they were abolished by low concentrations of atropine sulphate suggesting that the spontaneous contractions occurred as a result of acetylcholine released from cholinergic neurons to the mouse rectum. Similar observations were made in the present study. The effect of mefloquine when applied alone separately to the mouse rectum was relatively trivial and was variable. This effect of mefloquine on the mouse rectum differs from those of other antimalarials such as quinine observed in lizard and rat rectal muscles where this compound induced powerful activity (Savage and Akinialu, 1985, Savage and Lawal, 1986). The relatively weak agonist action of mefloquine may be indicative that the membrane of the mouse rectal smooth muscle may probably have a low resting membrane potential compared to those of the rat and lizard. When the extracellular Ca2+ concentration was increased, mefloquine induced pronounced rhythmic phasic contractile activity which was abolished on re-admission of normal (1.8mM Ca2+) Tyrode solution of Ca2+ free Tyrode solution. The contractile effects of mefloquine encountered under this condition were direct effects of this compound on the rectal smooth muscle and was not indirectly mediated by an enhanced release of acetylcholine from intra-mural cholinergic nerve fibers to the mouse rectum, as they were unaffected by atropine sulphate. This observation shows that the agonist action of mefloquine in this tissue is dependent not only upon the presence of extracellular Ca2+ but also on the Ca2+ concentration. It may be suggested that mefloquine can apparently only elicit contractile responses when the intracellular Ca2+ storage organelles are saturated. Mefloquine hydrochloride (4.1x10-5 – 5.2x10-3M) when applied alone caused variable responses of the mouse rectum. In some preparations no effect was recorded while in other preparation mefloquine produced very weak concentration-dependent contractions and slightly enhanced the rhythmic activity of the mouse rectum. However when the calcium concentration of bathing Tyrode solution (Ca2+) was increased from 1.8mM to 300mM, Mefloquine (4.1x10-5 – 5.2x10-3M) caused prominent contractile responses. Similar results were obtained when 1.4x10-6M of atropine sulphate was present in the bathing solution, suggesting that the mefloquine enhancement of contractile activity in high (Ca2+) is not mediated by acetylcholine release from either cholinergic neurons or the mouse muscle itself. Mefloquine concentration-dependently inhibited contractile responses induced by high potassium, carbachol or calcium in depolarizing Tyrode solution. Contractile responses induced by carbachol are brought about by stimulation of muscarinic cholinoceptors, since they were abolished by atropine sulphate. The carbachol and potassium chloride contractions appear to have occurred as a result of stimulation of extracellular Ca2+ since the contractile responses were reversible lost on Ca2+ - withdrawal. Carbachol-stimulated trans-membranal Ca2+-influx must have occurred via Ca2+ channels associated with muscarinic receptors – the so called receptor-operated Ca2+ Channels (ROC’S), whereas KCL-induced contractions are due to depolarizing action on the plasma-membrane of the mouse rectal smooth muscle as a result of which extracellular Ca2+ - influx occurs via Ca2+ channels in the smooth muscle (Haddart and price, 1976). The results of the experiments in this study show that mefloquine exhibits antagonist actions against various types of pharmacological interventions, which suggest that it is relatively unspecific in their antagonistic actions. This compound possesses appreciable Ca2+-antagonistic” and antimuscarinic activity at concentrations similar to plasma concentrations achieved during antimalarial therapy and may be responsible for some of the autonomic side effects encountered with the use of this compound in malaria (Unekwe et al, 1990). References
©Physiological Society of Nigeria, 2007 The following images related to this document are available:Photo images[np07007f1b.jpg] [np07007f1a.jpg] [np07007f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}